72
ИНДИВИДУАЛЬНО-ТИПОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ОРИЕНТИРОВОЧНОЙ РЕАКЦИИ В ПРОГНОЗИРОВАНИИ АДАПТАЦИИ ОПЕРАТОРОВ
Н.Р.
НЕСМЕЛОВА
Работа
выполнена при поддержке РГНФ, грант № 97 — 06 — 08133а.
Операторские профессии очень распространены в современном обществе.
Поэтому важное теоретическое и прикладное значение имеет изучение
психофизиологических особенностей человека, способных повлиять на результаты
операторской деятельности. Известно, что в основе обработки информации
оператором лежит ориентировочный рефлекс [12],
на характер которого влияют, среди других факторов,
индивидуально-типологические особенности человека: экстраверсия, нейротизм, особенности функциональной асимметрии и
вегетативной регуляции, а также свойство активированности
ЦНС [14], [16];
в структуре индивидуальности человека эти качества определенным образом
взаимосвязаны [9], что позволяет предполагать
существование типов индивидуальности, определяющих характер ориентировочной
реакции и успешность адаптации к операторской деятельности. Целью данной работы
явилась попытка выделить такие типы, используя кластерный анализ индивидуальных
психофизиологических особенностей человека [1],
а также изучение характера ориентировочного рефлекса и успешности адаптации к
операторской деятельности в зависимости от типа индивидуальности.
Экспериментальная часть исследования выполнялась в НИИ биологии и
биофизики при Томском государственном университете совместно с Ю.В. Бушовым. В экспериментах участвовали практически здоровые
мужчины в возрасте 18 — 36 лет, студенты. Всего обследовано 114 человек. В
программу обследования включалось выполнение теста Айзенка,
кистевая динамометрия, измерение слуховых порогов на частоте 1000 Гц, измерение
артериального давления и частоты сердечных сокращений (ЧСС). По результатам
кистевой динамометрии рассчитывался показатель моторной асимметрии как разность
между средними показателями правой и левой руки. По результатам измерения
слуховых порогов рассчитывался показатель сенсорной асимметрии как разность
между слуховыми порогами левого и правого уха. Результаты измерения частоты ЧСС
и давления использовались для вычисления вегетативного индекса Кердо. При исследовании ориентировочного рефлекса
испытуемый находился в звукоизолированной камере, в
положении сидя, с закрытыми глазами. В качестве источника звуковых стимулов
использовался фоностимулятор FNS-2. Звуковые тоны
интенсивностью 60 дБ, частотой 1000 Гц и длительностью 10 с (умеренный сигнал)
подавались через наушники
73
с межстимульными интервалами 20 — 30 с
до полного угасания кожно-гальванической реакции
(КГР), критерием которого служило отсутствие КГР при трех последовательных
сигналах. Число предъявлений стимула до полного угасания КГР использовалось для
оценки степени активированности ЦНС [8]. После 10-минутного перерыва однократно
предъявлялся звуковой сигнал интенсивностью 100 дБ, частотой 1000 Гц,
длительностью 10 с (сильный сигнал). С помощью полиграфа П8Ч-01
регистрировались электрокардиограмма (ЭКГ) в отведении А
по Небу, торакоспирограмма, реаграмма
на частоте 40 Гц с указательного пальца левой руки, КГР по Тарханову с
безымянного пальца левой руки испытуемого. При ручной обработке записей
измерялись длительность кардиоциклов, амплитуда зубца Т ЭКГ, период дыхания и длительность анакротической
части реографической волны в течение 10 с до подачи сигнала (фон), 10 с во
время действия сигнала и 20 с после его прекращения, а также показатели в
начале каждой секунды. Длительность кардиоциклов с
учетом скорости движения ленты переводилось в ЧСС, амплитуда зубца Т ЭКГ с учетом калибровочной записи по каждому испытуемому
представлялась в милливольтах, период дыхания с учетом скорости движения ленты
был представлен в секундах, длительность анакротической
части реографической волны в миллисекундах. Среднее
значение каждого показателя за 10 с перед
подачей сигнала определялось в качестве фона, значения в каждую секунду во
время и после действия стимула представлялось в процентах к соответствующему
фону.
В качестве модельной операторской деятельности
использовалась работа на компьютере с программой для изучения особенностей
восприятия времени "Hronos" [5]. Известно, что правильное ориентирование во
времени является важным фактором успешной работы оператора [2]. Испытуемому в случайном порядке предъявлялись
звуковые сигналы интенсивностью 60 дБ, частотой 1000 Гц, длительностью от 0,8
до 35,3 с. Прослушав сигнал, испытуемый должен был воспроизвести его длительность,
удерживая нажатой клавишу компьютера. При первом обследовании звук
каждой длительности предъявлялся пять раз. Для оценки качества работы
использовались средняя ошибка и дисперсия ошибки в процентах к реальной
длительности сигнала.
В опытах с трехдневной адаптацией к модельной операторской деятельности
участвовало 50 человек. Для исследования динамики обучения анализировались
значения средней ошибки воспроизведения звукового сигнала длительностью 1,3 с.
Ежедневно испытуемый дважды выполнял задание на воспроизведение длительности
звуков, контрольный сигнал (1,3 с) предъявлялся по 20 раз в каждом задании
среди сигналов других длительностей. В перерыве между заданиями (2 часа)
испытуемый выполнял какой-либо другой вид операторской деятельности (компенсаторное
слежение за синусоидальным сигналом или работа на компьютере, связанная с
решением арифметических примеров разной сложности) и психологические тесты.
Статистический анализ данных проводился с использованием программы "Statistica" [3].
Для выделения типов индивидуальности был проведен кластерный анализ набора
психофизиологических признаков, в который вошли экстраверсия и нейротизм по тесту Айзенка,
показатели моторной и сенсорной функциональной асимметрии, вегетативный индекс Кердо и показатель активированности
ЦНС. Для того, чтобы исключить признаки, связанные
между собой линейной связью, между отобранными для анализа показателями
предварительно были подсчитаны коэффициенты корреляции Спирмена,
которые оказались очень малы и не достигали 5 %-го уровня значимости.
Кластерный анализ проводился по алгоритму k-средних с использованием
евклидовой метрики [1].
74
В
анализ вошли данные по 109 из 114 участников опыта (результаты обследования 5
человек по разным причинам оказались неполными и были исключены). Оптимальное
число кластеров определялось методом перебора [1].
При анализе межгрупповых различий применялся критерий Манна - Уитни, а при
анализе динамики вегетативных компонентов ориентировочного рефлекса - критерий Вилкоксона. Считалось, что реакция имеет место, если
значение показателя в соответствующие периоды записи значимо отличалось от
фонового значения в данной группе. Различия считались достоверными при уровне
значимости p£0,05.
Таблица
|
Группы Признаки |
Группа 1 (N=23) |
Группа 2 (N=33) |
Группа 3 (N=21) |
Группа 4 (N=32) |
F |
p£ |
|
Э |
9,4±1,024 |
12,5±0,9134 |
10,1±1,124 |
14,2±0,8123 |
21,9 |
0,000 |
|
Н |
9,1±1,1234 |
11,3±1,0134 |
15,6±l,6124 |
12,9±1,3123 |
15,9 |
0,000 |
|
ВИ |
-9,8±8,84 |
-6,3 ± 5,84 |
-15,7±11,24 |
6,6±4,6123 |
5,1 |
0,002 |
|
МА |
6,7 ±1,623 |
3,3±1,3l34 |
-1,2±2,1124 |
5,8±1,123 |
19,2 |
0,000 |
|
СА |
5,2± 1,7234 |
2,0±1,51 |
0,7±1,81 |
1,7±1,21 |
6,7 |
0,000 |
|
А |
5,4±1,92 |
14,0±1,6134 |
6,8±4,724 |
3,2±1,923 |
66,3 |
0,000 |
Примечание. В таблице
приведены средние значения с 95 %-ми доверительными интервалами. Верхними
индексами отмечены достоверные межгрупповые различия, цифры соответствуют
номерам групп, при сравнении с которыми получено достоверное различие по
данному признаку. Сокращения: Э — экстраверсия (отн. ед.), Н — нейротизм (отн. ед.), ВИ — вегетативный индекс Кердо
(отн. ед.), МА — моторная асимметрия (кг), СА —
сенсорная асимметрия (дБ), А — активированность ЦНС (отн. ед.).
Было выбрано решение, состоящее из четырех кластеров, достоверно
различающихся между собой по всем использованным в анализе индивидуальным
признакам (см. табл.).
Межгрупповые различия в динамике вегетативных компонентов
ориентировочного рефлекса были обнаружены для всех исследованных компонентов,
большее количество значимых межгрупповых различий выявлено при анализе реакций
на умеренный сигнал. Различия по динамике ЧСС (рис. 1) представляют особый
интерес, поскольку изменения ЧСС в разные периоды после начала действия стимула
связывают с разными рефлексами (ориентировочный, оборонительный, стартл-рефлекс) [28].
В структуре сердечного компонента ориентировочного
рефлекса выделяют реакцию коротколатентного
замедления сердечного ритма (КЗСР) с латентным периодом (ЛП) от 0,5 до 2 сек. с
момента начала действия стимула, ускорение сердечного ритма (стартл-реакция) с ЛП до пика около 4 с, вторичное
замедление сердечного ритма (ЛП) от 2,5 до 13 с и длиннолатентные
изменения, которые могут быть представлены как увеличением, так и уменьшением ЧСС [22], [23], [27], [28]. В группе 1
(рис. 1 а) была зафиксирована достоверная реакция вторичного замедления
сердечного ритма и длиннолатентное уменьшение ЧСС. В
группе 2 имели место реакция КЗСР, вторичное замедление сердечного ритма и
незначительное, но достоверное длиннолатентное
повышение ЧСС (рис. 1 б). В группе 3 обнаружены
реакция КЗСР и длиннолатентное увеличение ЧСС (рис. 1
в). В группе 4 обнаружена только реакция вторичного уменьшения ЧСС (рис.
75
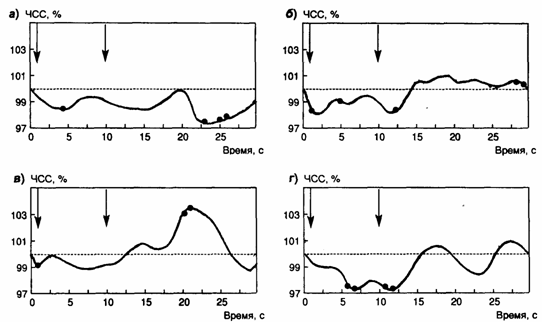
Рис. 1. Динамика ЧСС во время и после
действия умеренного сигнала в разных группах испытуемых. Стрелками показаны
моменты включения и выключения сигнала.
Отмечены достоверные отличия от среднефоновых
значений в каждой группе
При анализе успешности освоения операторской деятельности, связанной с
воспроизведением длительности звуковых сигналов, также выявлены межгрупповые
различия (рис. 2).
При первом обследовании группа 2 демонстрировала достоверно бóльшую ошибку воспроизведения контрольного сигнала
по сравнению с другими группами. В ходе последующих трех дней обучения
результаты во всех четырех группах первоначально улучшались, однако к
последнему дню опыта результаты группы 1 вернулись к исходному уровню, а
результаты группы 3 стали даже хуже, чем в первый день. В группах 2 и 4
достигнутое в первые дни обучения улучшение результатов сохранилось. В
последний день опыта результаты групп 1, 2 и 3 достоверно не различались между
собой, а результаты группы 4 оказались достоверно лучше, чем в любой другой
группе (см. рис. 2).
Анализ межгрупповых различий по психофизиологическим признакам (табл.)
показал, что группы, отличающиеся благоприятной динамикой адаптации (2 и 4),
имеют более высокие показатели экстраверсии по сравнению с группами, результаты
которых ухудшились в конце опыта (1 и 3). В литературе
представлены похожие данные, согласно которым при длительной работе интроверты
снижают уровень исполнения деятельности, связанной с обнаружением околопороговых сигналов, тогда как у экстравертов в тех же
условиях уровень деятельности продолжает улучшаться или ухудшается
незначительно [21]. Можно предполагать, что
худшая адаптация интровертов к длительной работе, связанной с восприятием и
анализом внешних сигналов, связана с особенностями ориентировочного рефлекса.
Анализ динамики сердечного компонента ориентировочной реакции при действии
умеренного сигнала показал, что в группах 1 (рис. 1 а) и 3 (рис. 1 в)
имеют место длиннолатентные изменения ЧСС.
Дисперсионный анализ позволил обнаружить
76
значимые
межгруповые различия по динамике ЧСС с 21 по 26 сек.
от момента включения умеренного сигнала (Rao R(18,274)=2,184,
p=0,004). Согласно предположениям зарубежных исследователей [23], [28], длиннолатентные изменения ЧСС связаны с оборонительным
типом реакции на внешний сигнал, однако в работах этих авторов использовались
звуки большой интенсивности (110 дБ и выше), которые заведомо должны были
вызвать оборонительный рефлекс у большинства испытуемых. В нашей работе
значительное число испытуемых (40 %) продемонстрировало подобные, хотя и менее
сильные реакции в ответ на звук умеренной интенсивности, который, казалось бы,
не должен вызывать оборонительного рефлекса. По данным Е.Н. Соколова [18], граница между ориентировочным и
оборонительным рефлексами на звуковые сигналы лежит в области 90 дБ. Однако ряд
исследователей, например П.В. Симонов [16],
указывают, что преимущественно ориентировочно-исследовательский или
оборонительный характер ориентировочного рефлекса зависит не только от
параметров сигнала, но и от индивидуально-типологических особенностей человека.
В то же время установлено [17], что для
интровертов характерна высокая активность миндалевидного комплекса, которая, по
некоторым данным [6], поддерживает
(усиливает) отрицательные эмоциональные реакции страха - ярости и тормозит
ориентировочно-исследовательскую активность. Кроме того, экспериментальные
данные свидетельствуют, что если субъекту предоставляется возможность самому
увеличивать освещенность и включать звуковой сигнал в камере, то интроверты, в
отличие от экстравертов, предпочитают большую часть времени находиться в
полумраке и в тишине [17]. Таким образом интроверты стремятся уменьшить действие внешних
стимулов, что как раз и составляет биологический смысл оборонительного рефлекса
[18]. Значит, уровень экстраверсии может быть
одним из индивидуально-типологических признаков, определяющих преобладание
ориентировочно-исследовательской или оборонительной тенденции в реагировании на
незнакомые сигналы.
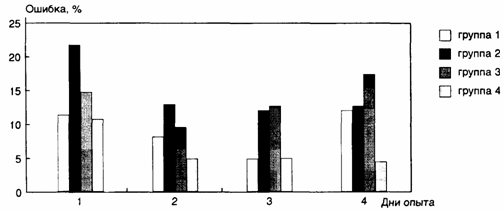
Рис.
2. Средние ошибки воспроизведения звуковых сигналов
длительностью 1,3 с
в
разных группах испытуемых в разные дни опыта
Разная направленность длиннолатентных изменений
ЧСС в 1 и 3 группах, возможно, свидетельствует о склонности к
активно-оборонительному (группа 3) или пассивно-оборонительному (группа 1)
поведению. Показано [25], что активно-агрессивные
субъекты отвечают на условный сигнал болевого раздражения учащением
сердцебиений, а пассивно-оборонительные - урежением.
77
Сравнение психофизиологических особенностей групп 1 и 3 (табл.)
показывает, что они резко различаются по уровню нейротизма
и по особенностям функциональной асимметрии. Для группы 1 характерно сочетание
низкого нейротизма с выраженным преобладанием правой
руки и правого уха, свидетельствующих о доминировании
левого полушария мозга. Для группы 3, напротив, характерно сочетание высокого нейротизма и преобладания левой руки, а показатель
сенсорной асимметрии в этой группе близок к нулю. Такой профиль функциональной
асимметрии свидетельствует об определенной степени доминирования правого
полушария либо об отсутствии доминирования. Низкий уровень нейротизма
предполагает высокую эмоциональную устойчивость и хороший контроль
за поведением, что связано со способностью быстро подавлять
"вегетативный взрыв", возникающий в эмоциогенных
условиях [11]. Установлено [10], что способность произвольно
расслабляться или дозировать степень напряжения утрачивается при поражениях
определенных отделов левого полушария. Анатомические данные
говорят о преимущественной связи левого полушария с активирующей ретикулярной
формацией ствола мозга (РФ) [4]. Вероятно, у
лиц с высокой степенью доминирования левого полушария (группа 1) при
возрастании под действием внешнего стимула возбуждающих влияний РФ на кору
срабатывает система отрицательной обратной связи, тормозящей активность РФ и
активные поведенческие реакции [13]. У лиц с
преобладанием правого полушария или без выраженной межполушарной
асимметрии эта система, вероятно, действует менее эффективно. Известно, что
возбуждение РФ является причиной ряда вегетативных сдвигов, характерных для
ориентировочной реакции, в частности, сужения периферических сосудов руки [15]. При сравнении степени сужения сосудов в разных
группах испытуемых выяснилось, что для группы 3 характерны максимальные, а для
группы 1 - минимальные по величине и длительности сосудистые реакции. Изменение
периферического кровотока [22] при использовании
умеренного сигнала в группе 3 составило 19,5±2,9 %, а в группе 1 — 11,0±2,3 % (p<0,01). Таким образом, можно
предполагать, что повышенная эффективность "системы торможения
поведения" [13] у лиц с выраженным
доминированием левого полушария (группа 1) способствует реализации
пассивно-оборонительного типа реагирования, заключающегося в уменьшении воздействия
внешнего сигнала на организм путем развития охранительного торможения в ЦНС,
повышения сенсорных порогов и подавления вегетативных реакций, а сниженная
эффективность этой системы при доминировании правого полушария способствует
активным ответам ("борьба", т.е. попытки устранить источник
стимуляции, или "бегство" от него), для энергетического обеспечения
которых необходимы значительные вегетативные изменения. Известно,
что особенности функциональной асимметрии могут способствовать возникновению психовегетативной дезинтеграции, для которой характерны
неадекватные (повышенные или сниженные) вегетативные реакции на внешнюю
стимуляцию и неадекватное вегетативное обеспечение различных видов
деятельности, что приводит к быстрому развитию утомления [7]. По-видимому, именно утомление явилось основной
причиной снижения результатов работы в последние дни опыта в группах с
неблагоприятной динамикой адаптации.
Группы, продемонстрировавшие благоприятную динамику адаптации к операторской
деятельности (1 и 3), отличаются более высокой экстраверсией, средним уровнем нейротизма и умеренной степенью правосторонней
межполушарной асимметрии (табл.). Для них характерно также отсутствие
выраженных длиннолатентных изменений ЧСС. Однако результаты
операторской деятельности в этих группах различны: величина ошибки
воспроизведения длительности
78
контрольного сигнала в группе 2 оказалась достоверно больше, чем в группе
4, как в первый, так и в последний дни опыта. При
сравнении психофизиологических особенностей этих групп (табл.) видно, что
наиболее резко они различаются по степени активированности
ЦНС. При сравнении динамики ЧСС в группах 2 (рис. 1 б) и 4 (рис.
Таким образом, наиболее перспективной группой в плане адаптации к
операторской деятельности оказалась группа 4. Для представителей этой группы
характерны самые высокие значения экстраверсии, средние уровни нейротизма и функциональной асимметрии, положительные
значения вегетативного индекса Кердо,
свидетельствующие о преобладании тонуса симпатического отдела вегетативной
нервной системы (табл.). Важность учета этих качеств при
профессиональном отборе будущих операторов подтверждается анализом литературы ([12], [14] и
др.).
В качестве дополнительной проверки полученных данных были
проанализированы результаты компенсаторного слежения за синусоидальным
сигналом,
79
которое
выполняли некоторые испытуемые в течение трех дней по два часа в день в
перерывах между первым и вторым сеансами работы с программой "Hronos". Из-за небольшого числа испытуемых анализ проводился
путем сравнения индивидуальных кривых динамики обучения наиболее типичных
представителей разных групп, отобранных с учетом индивидуальных расстояний от
кластерных центров.
Оказалось, что представитель группы 4 уже во втором часу первого дня обучения
значительно уменьшил число "аварий", которые регистрировались при
возникновении ошибки слежения выше допустимого уровня и сопровождались
аварийным звуковым сигналом. Во второй и третий дни обучения этот испытуемый
сохранял высокое качество работы. Представитель группы 2 также
продемонстрировал благоприятную динамику обучения, однако улучшение его
результатов происходило значительно медленнее. Представитель группы 3 во второй
день несколько улучшил свои результаты, а в третий день вновь ухудшил. Представителей
группы 1 среди испытуемых, выполнявших компенсаторное слежение, не оказалось.
Таким образом, анализ результатов компенсаторного слежения показал те же
закономерности обучения в группах, что и анализ результатов воспроизведения
длительности звуковых сигналов.
1. Выделено четыре группы испытуемых, различающихся по ряду
индивидуальных психофизиологических особенностей, по характеру вегетативных
компонентов ориентировочного рефлекса и по динамике адаптации к разным видам
операторской деятельности.
2. Показано, что для успешной адаптации к операторской
деятельности важны высокий уровень экстраверсии, преобладание влияний
симпатического отдела вегетативной нервной системы, низкая активированность
ЦНС, средний уровень нейротизма и умеренная степень
правосторонней функциональной асимметрии.
3. Показана перспективность изучения вегетативных компонентов
ориентировочной реакции для прогноза успешности адаптации к операторской
деятельности, а также для разработки основанных на непроизвольных реакциях
способов психодиагностики некоторых особенностей индивидуальности.
1. Айвазян С.А. и соавт. Классификация и снижение размерности. М.: Финансы и
статистика, 1989.
2. Батуев А.С. Высшая нервная
деятельность. М.: Высшая школа, 1991.
3. Боровиков В.П.,
Боровиков И.П. Statistica. Статистический анализ
и обработка данных в среде Windows. М.: Информационно-изд. дом "Филин", 1997.
4. Брагина Н.Н., Доброхотова Т.А. Функциональные асимметрии человека.
М.: Медицина, 1988.
5. Бушов
Ю.В., Несмелова Н.Н. Индивидуальные особенности восприятия человеком
длительности интервалов времени // Физиология человека. 1994. Т. 20. № 3. С.
30.
6. Вайнштейн И.И., Симонов
П.В. Эмоциогенные структуры мозга и сердце. М.:
Наука, 1979.
7. Вейн А.М.,
Соловьева А.Д. Лимбико-ретикулярный комплекс и вегетативная регуляция. М.:
Наука, 1973.
8. Голубева
Э.А. Способности и индивидуальность. М.: Прометей, 1993.
9. Голубева
Э.А. Структура индивидуальности в аспекте единства природного и социального
факторов // Природа психического: Тезисы докладов. Пермь, 1994. С. 66.
10. Доброхотова
Т.А., Брагина Н.Н. Функциональная асимметрия и психопатология основных
поражений мозга. М.: Медицина, 1977.
11. Изюмова
С.А., Аминов Г.А. О физиологической природе связей между эмоциональной
устойчивостью и свойствами нервной системы человека // Вопр.
психол. 1978. № 5. С. 128 — 133.
12. Крылов А.А.
Организация целостной деятельности функциональных механизмов обработки
информации // Хрестоматия по инженерной психологии. М.: Высш.
школа, 1991. С. 158 — 170.
13. Мартон М.
Типология и активация // Психология личности в социалистическом обществе. М.:
Наука, 1990. С. 47 — 60.
14. Несмелова Н.Н., Бушов Ю.В. Влияние индивидуальных психофизиологических
особенностей человека на характер ориентировочного рефлекса // Сиб. психол. журн. 1996. Вып. 2.
С. 67.
15. Ройер Л. Вегетативные
компоненты ориентировочного рефлекса // Нейронные механизмы ориентировочного
рефлекса. М.: Изд-во МГУ, 1970. С. 397 — 404.
16. Симонов П.В.
Высшая нервная деятельность человека. Мотивационно-эмоциональные аспекты. М.:
Наука, 1975.
17. Симонов П.В.
Мотивационный мозг. М.: Наука, 1987.
18. Соколов Е.Н.
Восприятие и условный рефлекс. М.: Изд-во МГУ, 1958.
19. Соколов
Е.Н. Нейронные механизмы памяти и обучения. М.: Наука, 1981.
20. Тихомирова И.В.
Формы активации и особенности стиля познавательной деятельности // Психология и
психофизиология индивидуальных различий в активности и саморегуляции поведения
человека. Свердловск, 1985. С. 49 — 54.
21. Фришман
Е.З. Вариативность порогов обнаружения и ее
причины // Психофизика сенсорных систем. М.: Наука,
1979. С. 46 —
63.
22. Barry
R.J. Novelty and significance effects in the fractionation of phasic OR measures: A synthesis with traditional OR theory
// Psychophysiol. 1982. V. 19. N 1.
P. 28 — 35.
23. Eves
F.F., Gruzelier J.H. Individual differences in
the cardiac response to high intensity auditory stimulation // Psychophysiol. 1984. V. 21. N 3. P. 342 — 352.
24. Furnham
A., Gunter B., Peterson E. Television distraction and the performance of
introverts and extroverts // Appl. Cogn. Psychol. 1994. V. 8. N 7. P. 705 — 711.
25. Hein
P., Cohen S., Shmavonian B. Perceptual mode and
cardiac conditioning // Psychophysiol. 1966. V. 3. N 2. P. 101 — 107.
26. Lacey J. et all. The visceral level: Situational determinants and behavioral correlates of autonomic response patterns // Expression of emotions in man. N.Y., 1963. P. 161 — 205.
27. Raine A., Venables
P.H., Williams M. Autonomic orienting response in 15-years-old male
subjects and criminal behaviour at age 24 // Am. J.
Psychiatry. 1990. V. 147. Iss. 7. P. 933 — 937.
28. Turpin G. Effects of stimulus intensity on autonomic responding: The problem of differentiating, orienting and defence reflexes // Psychophisiol. 1986. V. 23. N 1. P. 1 — 14.
Поступила
в редакцию 22.IV