143
МЕХАНИЗМЫ АДАПТАЦИИ К
СТИМУЛУ ВЫСОКОЙ ИНТЕНСИВНОСТИ
Э. М. РУТМАН, Б. И. КОЧУБЕЙ
Изучение психофизиологических механизмов адаптации к
стрессу — необходимый этап решения актуальнейшей современной проблемы повышения
стрессоустойчивости человека. В экспериментальных
исследованиях этого направления, наряду с болевой электрокожной
или холодовой стимуляцией, часто применяются в
качестве стрессора стимулы высокой интенсивности. До недавнего времени об
индивидуальных особенностях реакции на стресс и стрессозащитных
механизмах судили по изменениям показателей вегетативной нервной системы (ЧСС,
КГР и т.п.), реже — ЭЭГ и лишь сравнительно недавно стали использовать
вызванные потенциалы (ВП). Между тем многократно описанный феномен снижения
амплитуды определенных компонентов ВП при повышении интенсивности стимула,
начиная с некоторого индивидуально различного уровня [15],
позволяет предположить, что в ВП отражается активность некоторого стрессозащитного механизма, направленного на снижение
эффектов чересчур интенсивной стимуляции. В частности, можно ожидать, что
характеристики привыкания ВП, коррелирующие, по имеющимся данным, с рядом
психологических характеристик [18], окажутся ценными
индикаторами индивидуальных особенностей адаптации к стрессу и инструментом
исследования их механизмов.
В данной работе представлены
результаты исследования привыкания компонентов ВП при повторении тонов двух
интенсивностей, одна из которых, по данным литературы, вызывает ориентировочную
реакцию (ОР), другая — оборонительную [8], [21].
Анализ разноречивых данных о связи привыкания с интенсивностью стимула ([21], [27] и др.) привел нас к
предположению, что изменение характеристик привыкания при повышении
интенсивности может быть обусловлено хотя бы отчасти появлением оборонительной
реакции и включением связанных с ней защитных механизмов, направленных на
снижение эффектов чрезмерной интенсивности. Поскольку, по многочисленным
данным, увеличение частоты сердечных сокращений (ЧСС) является одним из
признаков оборонительной реакции, тогда как ОР характеризуется, напротив, урежением сердечного ритма (обзор см. [21]),
мы выделили при предъявлении громкого тона группу лиц с достоверным увеличением
ЧСС и провели, в частности, сопоставление характеристик привыкания у лиц с
учащением ритма и с отсутствием такового.
Поскольку данная работа
является частью более широкого исследования, посвященного выявлению соотношения
генетических и средовых факторов в формировании индивидуальных особенностей
привыкания, она проведена на близнецах.
МЕТОДИКА
Испытуемыми были 22 пары
моно- и 21 пара дизиготных близнецов в возрасте 17—
144
29 лет. Среди монозиготных пар было 10 мужских и 12
женских, среди дизиготных — 9 мужских и 12 женских.
Во время опыта испытуемые сидели в удобном кресле в темной звукоизолированной
камере. После периода адаптации (около 2 мин) без предупреждения предъявляли 33
тона 1000 Гц, 80 дБ над уровнем 0,0002 бара длительностью 480 мс с межстимульными интервалами 10 с. После 30-секундной паузы
начиналась другая серия из 33 тонов, отличающихся только интенсивностью,
которая была равна 105 дБ. Предварительно испытуемым говорили, что возможные в
связи с проверкой приборов внешние стимулы не требуют от них никаких реакций.
Источником звуков служил звукогенератор ЗГ-31, звуки
предъявлялись через динамик, расположенный на расстоянии 1 м спереди от
испытуемого и скрытый от его глаз.
ЭЭГ регистрировали монополярно в точке С с
объединенным ушным электродом в качестве индифферентного. Электроокулограмму
(ЭОГ) регистрировали также монополярно, с активным
электродом над наружным углом левого глаза. Специально разработанная методика [5] применялась для устранения влияния мигательных артефактов.
Для регистрации ЧСС использовали электрод на левом предплечье и ушной электрод.
Запись электрической активности производили на электроэнцефалографе
«Нихон Коден» и
магнитографе ТЕАС с постоянной времени 0,1 с (для — ЧСС — 0,03 с); фильтр
высоких частот — 60 Гц. На ЭВМ АТАС-350 усредняли ВП с периодом анализа 2 с
(включая 0,5 с до начала стимула) и шагом дискредитации 4 мс. Усреднение
производилось по 10 одиночным ВП: 1 — 10-му, 4—13-му, 11—20-му, 21—30-му;
впоследствии эти ВП обозначаются как ВП1—10, ВП4—13 и т.
д. Цель исследования — выявление динамики ВП при повторении стимула — диктовала
необходимость относительно небольшого числа усреднений, как это делается во
всех работах такого рода [19], [24].
Полученные ВП выводились на графопостроитель для последующего визуального
анализа.
РЕЗУЛЬТАТЫ
1. Описание ВП. В соответствии
с характеристиками большинства ВП было выделено 6 компонентов, обозначаемых
буквой, соответствующей полярности, и порядковым номером: Р1, N1, Р2, N2, Р3,
N3. Названные компоненты имели латентные периоды до пика 30—75 мс, 85—140 мс,
160—220 мс, 215—280 мс, 245—400 мс и 335—500 мс соответственно. Амплитуда
компонентов определялась от средней линии, для получения которой находили
среднее значение между максимумом и минимумом амплитуды на отрезке 0,5 с до
начала стимула. В некоторых ВП (19 %) компоненты N2 и Р3 отсутствовали, и Р2
непосредственно переходил в N3. В таких случаях максимальная негативность в
интервале 230—280 мс принималась за амплитуду N2, а максимальная позитивность
на отрезке 250—350 мс — за амплитуду Р3.
2. Общие характеристики
привыкания. Зависимость ВП от интенсивности и привыкания изучалась с
помощью двухфакторного дисперсионного анализа 2 (интенсивность) × 4
(привыкание). При обнаружении значимых F-отношений проводилось попарное сравнение средних и других контрастов по критерию Тьюки [2].
Для латентных периодов (ЛП)
большинства компонентов не обнаружено значимых изменений ни в связи с
интенсивностью, ни по мере повторения стимулов. Только ЛП компонента Р3
укорачивается при повышении интенсивности (F=14,33; p<5%); при повторении стимула изменений этого показателя не
отмечается. Повышение интенсивности вызывает значимое увеличение амплитуд
компонентов P1 (F= 17,23; p<0,1%), N1 (F=182,84; p<0,01%), Р2 (F=50,81; p<0,01 %) и Р3 (F=37,22; p<0,01%). Амплитуда N2 при увеличении интенсивности уменьшается (F=74,86,
p<0,01%). Амплитуда N3 от интенсивности
существенно не зависит.
Значимое снижение амплитуды
при повторении стимулов (привыкание) обнаружено только для компонентов N1 (F=
17,04, с p<0,1%) и Р3 (F=4,38; p<1%). Методом попарных сравнений выявлено
много значимых контрастов для компонента N1. Так, можно отметить весьма
«крутое» начало привыкания этого компонента при повторении тона 105 дБ: только
при этой интенсивности значима разница между ВП1—10 и ВП4—13.
При 80 дБ, напротив, снижение амплитуды больше выражено в конце серии стимулов:
так, при обеих интенсивностях имеется значимая разница между ВП1—20
и ВП21—30, но только при 80 дБ отсутствуют различия между ВП1—10
и ВП11—20. Для Р3 при обеих интенсивностях достоверны только
контрасты между ВП1—10 и ВП21—30. а также между ВП1—10
и ВП11—30. Амплитуда N2 увеличивалась при повторении стимула, это
увеличение достигало достоверности лишь для стимуляции 80 дБ при сравнении ВП21—30
с ВП1—10 и ВП4—13.
145
Таблица 1
Амплитуда и скорость
привыкания компонентов ВП в целом по группе 86 человек
|
Интенсивность |
Компоненты |
|||||
|
Р1 |
N1 |
Р2 |
N2 |
Р3 |
N3 |
|
|
Амплитуда, мкВ |
||||||
|
80 дБ 105 дБ |
3,0±0,52 4,7±0,56 |
27,4±1,27 44,0±2,08 |
22,7±1,35 28,9±1,26 |
—2,7±0,97 —7,4±1,07 |
11,1±1,25 16,1±1,40 |
12,5±0,84 12,4±0,81 |
|
Привыкание |
||||||
|
80 дБ 105 дБ |
—0,05+0,03 0,00+0,04 |
—0,35±0,05 —0,58±0,06 |
—0,18±0,05 —0,06±0,06 |
0,22±0,05 0,09±0,05 |
—0,23±0,05 —0,22±0,06 |
—0,04±0,04 —0,03±0,05 |
Примечание: амплитуды ВП даны по ВП1—10.
Для сопоставления с данными
других авторов, измерявших амплитуду от пика до пика, мы проанализировали
изменения амплитуд Р1 —N1 и N1—Р2. Амплитуда P1 —N1 увеличивалась с увеличением
интенсивности (F= 206,90; p<0,01%) и уменьшалась при
повторении (F= 13,26; p<0,1%). При обеих
интенсивностях достоверны различия по этому компоненту между ВП1—10 и ВП21—30, а при 105 дБ — также между ВП1—10 и ВП11—20, ВП1—10 и ВП4—30, ВП4—13 и ВП21—30. Амплитуда Р1—Р2 также
увеличивалась при повышении интенсивности (F= 164,48; p<0,01%) и уменьшалась при повторении (F=9,51; p<1%). При обеих интенсивностях ВП21—30 меньше ВП1—10, ВП4—13 и ВП1—20.
Для нахождения параметров
привыкания были рассчитаны коэффициенты линейной регрессии амплитуд компонентов
ВП по среднему номеру стимулов, взятых для усреднения, а для коэффициентов с
выраженным привыканием (N1 и Р3) — также коэффициенты регрессии по логарифму не
мера стимула. Результаты представлены в табл. 1 и на рис. 1.
Сравнение линейной и логарифмической
аппроксимаций выявило два случая достоверных различий: для привыкания Р1—N1 при
105 дБ ошибка логарифмической аппроксимации меньше ошибки линейной (F=12,00; df=3,3; p<5%). Привыкание N1—Р2 при
стимуляции 80 дБ, наоборот, достоверно лучше описывается линейной зависимостью (F= 10,87; df=3,3; p<5 %)
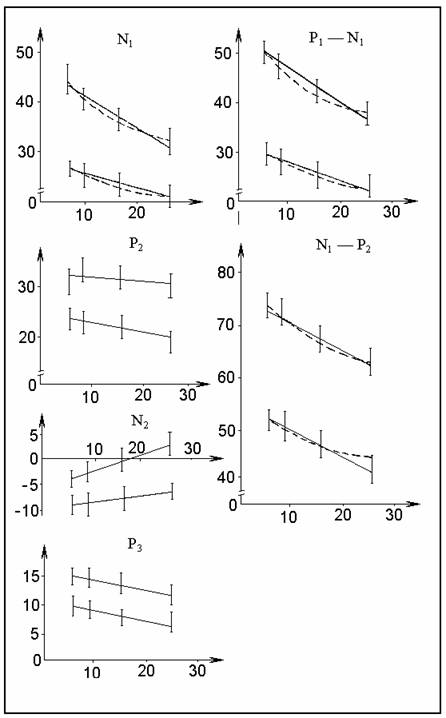
Рис.
1. Динамика
изменений амплитуды компонентов ВП при повторении стимула. Ось абсцисс — номер
стимула, ось ординат — амплитуда,
мкв. Вверху — 105 дБ, внизу — 80 дБ. Сплошная линия —
линейная регрессия, прерывистая —
логарифмическая. Компоненты обозначены
на рисунке.
С целью дополнительной
проверки равномерности хода привыкания компонентов ВП разность между
амплитудами ВП1—10
и ВП11—20 сравнивалась с разностью
между ВП11—20 и ВП21—30. Для компонента N1 при
146
интенсивности 105 дБ первая разность значимо
превышает вторую (t=3,73; p<1%), в остальных случаях
различий не было.
Таблица 2
Интеркорреляции между показателями
привыкания ВП на тоны 80 дБ (выше диагонали) и 105 дБ (ниже диагонали)
|
Компоненты |
№ 1 |
Р2 |
№2 |
Р3 |
|
N1 |
X |
23* |
08 |
—06 |
|
Р2 |
—35** |
X |
—24* |
27* |
|
N2 |
20 |
—61** |
X |
—62** |
|
Р3 |
16 |
13 |
—32** |
X |
Примечание. * — p<5 %, ** — p<1 %. Нули
и запятые опущены.
3. Межиндивидуальные
корреляции показателей привыкания. Ни для одного из компонентов ВП не
обнаружено значимых корреляций скорости привыкания (по коэффициенту линейной
регрессии) при повторении тона 80 дБ со скоростью привыкания того же или
другого компонента при повторении звука 105 дБ. Коэффициенты корреляции между
привыканием разных компонентов при одной и той же интенсивности приведены в
табл. 2.
4. Особенности привыкания
ВП на интенсивный стимул у лиц с наиболее выраженной оборонительной реакцией. По
величине ЧСС было выделено 36 испытуемых, у каждого из которых средняя ЧСС при
стимуляции тоном 105 дБ достоверно (по t-критерию) превосходила
среднюю ЧСС покоя и (или) ЧСС при стимуляции 80 дБ. Оказалось, что лица с
достоверным увеличением ЧСС при тоне 105 дБ отличаются от остальных более
быстрым привыканием N1 (t =2,02; p<5 %) и уменьшением привыкания Р2 (t=2,27; p<5 %) и Р3 (t=4,40; p<0,1 %). Точнее говоря, у этих испытуемых наблюдалось увеличение, а
не снижение амплитуды компонентов Р2 и Р3 при повторении тона 105 дБ (рис. 2).
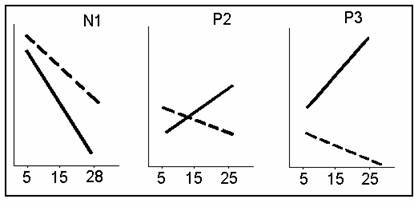
Рис.
2.
Динамика изменений компонентов ВП при повторении тона 105 дБ у испытуемых с
наиболее выраженной ОбР (сплошная линия) и менее
выраженной ОбР (пунктирная линия).
ОБСУЖДЕНИЕ
Полученные характеристики
привыкания при интенсивности 80 дБ соответствуют преобладающим в литературе
данным: достоверно снижается амплитуда компонентов N1 и Р3 [18],
[19], [24], [27].
Отсутствие значимого уменьшения компонентов Р1 и Р2 позволяет предположить, что
снижение амплитуд Р1 — N1 и N1—Р2, полученное в данном исследовании и часто
отмечающееся в литературе, обусловлено преимущественно привыканием компонента
N1. (Неожиданное уменьшение компонента N2 при увеличении интенсивности, его
увеличение при повторении в сочетании с сильными отрицательными корреляциями
привыкания N2 с привыканием Р2 и Р3 объясняются, на наш взгляд,
«несамостоятельностью» компонента N2 в том смысле, что его динамика может пассивно
отражать динамику соседних положительных компонентов. Сходные предположения
высказывались в литературе [29].)
В свете существующих
представлений об отражении в амплитуде N1 и Р3 величины рассогласования между
ожидаемым и реальным событиями [7], [25]
полученный результат свидетельствует, что основным механизмом наблюдавшегося
привыкания было переформирование прогноза (создание новой, адекватной «нервной
модели стимула» [9]) при повторении стимула и,
соответственно, уменьшение рассогласования, как это предполагается в широко
известной теории Е.Н. Соколова [9]. Этому вполне
соответствует и факт положительной корреляции привыкания компонентов N1, Р2 и
Р3 при повторении тона 80 дБ.
При переходе к интенсивности
105 дБ отмечается ряд существенных изменений. Во-первых, увеличивается
«крутизна» привыкания компонента N1: только в этом случае привыкание лучше
описывается «крутой» логарифмической зависимостью, чем «плавной» линейной, а
снижение амплитуды в первой половине серии значимо превышает снижение во
второй. Во-вторых, при повторении тона 105 дБ между привыканием N1 и Р2
наблюдается отрицательная корреляция, тогда как при тоне 80 дБ отмечалась
положительная. Выше мы замечали, что ни для одного компонента не обнаружено
значимых корреляций привыкания при тонах разной интенсивности. Добавим к этому,
что при переходе от интенсивности 80 дБ
147
к 105 дБ меняется соотношение гено-
и паратипических влияний на привыкание, как показало
специальное исследование, проведенное на этой же выборке близнецов [3].
Все перечисленные столь существенные изменения характеристик привыкания
компонентов ВП при переходе от 80 к 105 дБ не кажутся неожиданными в свете
существующих представлений о разной природе «ориентировочной» и
«оборонительной» активации [28], [30].
Эти представления, кстати, существенно подкрепляются тем фактом, что привыкание
компонента Р2, отражающего уровень активации [20], оказалось
значительно детерминировано генетическими факторами только при повторении тона
105 дБ [3].
Рассмотрим возможные
механизмы различных изменений привыкания разных компонентов ВП при увеличении
интенсивности.
Увеличение привыкания N1 не
представляется неожиданным. Во-первых, в литературе неоднократно ([11],
[21]) отмечалось, что предположение о прямой связи
привыкания с интенсивностью [31] эмпирически довольно часто
не подтверждается, более того, есть немало данных об увеличении привыкания с
увеличением интенсивности стимула [27]. Во-вторых,
многочисленные данные ([10], [13], [15], [16]) свидетельствуют о существовании
в нервной системе защитного механизма, направленного на снижение чересчур
высокого уровня активации до оптимального. Этот механизм мыслится либо как
некое подобие «сенсорного фильтра», снижающего величину реакций сенсорных
систем и одновременно субъективную величину интенсивности стимула ([15], [16]), либо как
модально-неспецифический «деактивирующий» механизм [10], [13]. Ускорение привыкания N1 при повышении интенсивности, особенно
ярко выраженное при первых предъявлениях стимула (см. табл. 1), вполне
соответствует этим представлениям.
Однако все же остается
неясным, почему возникает отрицательная корреляция между привыканием N1 и Р2
при предъявлении тона 105 дБ. Из вышеупомянутых представлений не следует также
и обнаруженный нами факт увеличения амплитуды Р2 и Р3 при повторении стимула
105 дБ у лиц с отчетливой оборонительной реакцией на этот стимул при
одновременном усилении у них привыкания N1 (см. рис. 2). Можно ли на сегодня
как-то понять эту совокупность фактов? Нам показалось правдоподобным следующее
предположение.
Привыкание можно
рассматривать как преодоление конфликта между воспринятым внешним событием и
его априорной репрезентацией в нервной системе. Этот конфликт можно преодолеть,
разрешить, изменяя либо реальность (внешнюю реальность или ее субъективные
характеристики), либо свое априорное представление о ней («нервную модель»,
«вероятностный прогноз» или «образ потребного будущего» — разные термины в
разных концепциях используются для обозначения разных сторон одного и того же
явления). Экспериментальные исследования ОР проводились, как правило, в
ситуациях, когда внешние изменения незначительны и несущественны (ОР
традиционно понимается как реакция на индифферентный стимул). В таких
условиях по мере повторения неожиданного (сначала) стимула происходит
ассимиляция нового опыта (изменение «нервной модели стимула», переформирование
«вероятностного прогноза», изменение «образа потребного будущего» — в
зависимости от концепции, в рамках которой рассматривается ситуация) и
восстанавливается соответствие, согласованность ожидаемых и реальных событий.
Подчеркнем, что при этом ожидаемые события не требуют изменений наличной
запрограммированной активности, которая может быть так называемым «покоем»
(наиболее частая экспериментальная ситуация исследования ОР), а также какой-то
деятельностью разной сложности. В противном случае либо говорят о возникновении
ОР на сигнальный стимул, либо рассматривают возникающие изменения в контексте
внимания, где опять-таки используются понятия типа «ожидания», «эталона»,
«сличения» и т.п., близкие к ранее названным.
В концепции «нервной модели
стимула», в русле которой главным образом рассматриваются явления привыкания ОР
у человека, имплицитно содержится представление о «точном» отражении физических
характеристик стимула в том паттерне нервной активности, который вызывается
стимулом и сличается с «нервной моделью». В рамках этой концепции не
рассматривается влияние на величину рассогласования тех факторов, которые в
психофизике, например в теории обнаружения сигнала, объединяются под названием внесенсорных и включают такие, в частности, факторы, как
степень предубежденности, значимость и т.п.
Между тем мы полагаем, что
подобно тому, как субъект при высоком уровне предубежденности (низком критерии)
при обнаружении сигнала воспринимает сигнал там, где его на самом деле нет, он
может не воспринять событие или рассогласование
148
между ожиданием и реальным событием там, где оно на
самом деле есть. Иначе говоря, он может не воспринять стимул там, где налицо
физическое действие стимула.
Так же как и в концепции
«нервной модели стимула», в исследованиях привыкания не рассматривалась
возможность устранения рассогласования при повторении стимула за счет субъективного
изменения восприятия в соответствии с «нервной моделью» или «образом потребного
будущего» для избежания рассогласования там, где оно является для индивида
нежелательным, а другие способы его устранения невозможны или трудны. Между тем
психологический феномен «принятия желаемого за действительное» хорошо известен
в жизни, многократно описан в художественной литературе, исследовался в рамках
психологии восприятия, в частности в работах так называемого нового взгляда [1], [14].
Мы предполагаем, что при
действии громкого звука у тех испытуемых, для которых звук настолько неприятен,
что «смириться» с ним (перестроить прогноз, как в случае тона 80 дБ) и
вернуться к предшествовавшему состоянию «покоя» они не могут, а изменить
внешнюю реальность — прервать опыт — также не могут из-за установки на
сотрудничество с экспериментатором, возникает явление, родственное так
называемой перцептивной защите [14]:
испытуемые начинают слышать звук как менее громкий, как не отличающийся или
мало отличающийся от ранее предъявлявшегося индифферентного тона.
Установлено, что амплитуда
N1 соответствует субъективной и «желательной» оценке интенсивности стимула: она
тем выше, чем выше человек оценивает интенсивность и чем более интенсивный
стимул он хотел бы услышать [6], [12].
Ускорение снижения амплитуды
N1 у лиц с увеличением ЧСС может быть отчасти отражением снижения субъективной
интенсивности громкого звука, возникающего в силу стремления испытуемых
игнорировать его неприятные свойства, «слышать» звук не слишком громким. Это, в
свою очередь, обеспечит устранение рассогласования и дополнительное снижение
амплитуды N1. Такому предположению соответствует тот факт, что у лиц с
наибольшим увеличением ЧСС (за небольшим исключением) отмечалась часто оценка
звука 105 дБ как не обладающего неприятными свойствами, а лица с небольшими сдвигами
ЧСС считали его весьма неприятным [4].
Возникает вопрос: почему
уменьшение N1, означающее, по нашему мнению, снижение субъективной
интенсивности и восстановление согласования, не сопровождается снижением
амплитуды Р2, связанной с уровнем активации, и Р3, отражающей степень
рассогласования ожиданий и реальности?
По имеющимся в литературе
данным, компоненты N1 и Р3 отражают рассогласование на разных уровнях: N1
преимущественно связан с уровнем, который можно условно назвать перцептивным, а
Р3 — с более обобщенным уровнем, включающим ожидания не только перцептивных
характеристик, но и требуемых моторных реакций и собственного состояния,
которое в значительной степени определяется вегетативными сдвигами. У лиц,
достигающих устранения рассогласования на перцептивном уровне за счет
иллюзорного восприятия пониженной интенсивности, не происходит адекватной
реорганизации всей совокупности ожиданий, в частности это относится к уровню
активации и вегетативных сдвигов, вызываемых громким звуком. В результате
многие явления, связанные с неприятной ситуацией, продолжают оставаться
неожиданными, уровень активации и рассогласования (отражаемого в Р3) не
снижается и может даже повышается, поскольку ожидание восстановления «покоя»,
которое наступило после предъявления какого-то числа тонов 80 дБ, все больше
расходится с действительностью при повторении тонов 105 дБ. Мы видим в этом
случае как бы плату за психическую защиту такого типа, плату, которая не
является неожиданной: известно, в частности, что перцептивная
защита сочетается с высоким уровнем вегетативной и корковой активации [17], [22]. По-видимому, стратегия,
отражающаяся в снижении N1, не приводит к вполне эффективной адаптации, так как
другие физиологические сдвиги — ЧСС, компоненты Р2 и Р3 ВП — при этом не
уменьшаются, а увеличиваются.
Нам кажется, что
обнаруженный феномен «диссоциации» изменений компонентов N1, Р2 и Р3 при
действии громкого звука может представлять собой явление, аналогичное
диссоциации разных показателей болевых ощущений при гипнотической аналгезии. У
лиц в состоянии гипнотической аналгезии исчезают болевые гримасы, но
сохраняются вегетативные реакции на болевые раздражители. Вербально болевое
воздействие отрицается, но при особых приемах, позволяющих отмечать восприятие
боли, не нарушая «соглашения» с гипнотизером, обнаруживается положительный
ответ на вопрос о боли [23]. Эта аналогия приводит
149
к мысли, что как гипнотическая аналгезия, так и
снижение субъективной неприятности громкого звука могут обеспечиваться одним и
тем же механизмом. Мы осмелились высказать это предположение, поскольку оно
доступно простой экспериментальной проверке. Если оно верно, то при
гипнотической аналгезии должно наблюдаться снижение амплитуды N1 при
одновременном повышении амплитуд Р2 и Р3 по сравнению с теми же компонентами ВП
на тот же болевой стимул без аналгезии.
ЗАКЛЮЧЕНИЕ
С помощью метода вызванных потенциалов
исследовались психофизиологические механизмы привыкания к индифферентному и
неприятному раздражителям. Обнаружено, что характеристики привыкания различны в
зависимости от наличия или отсутствия качества неприятности стимула, а также от
выраженности оборонительной реакции на неприятный стимул. Предполагается, что
индивидуальные различия в привыкании к нейтральной стимуляции связаны
преимущественно со скоростью перестройки «нервной модели стимула». При переходе
к неприятной стимуляции у ряда лиц возможно включение дополнительных механизмов
привыкания, связанных с регуляцией субъективно воспринимаемых характеристик
стимула, что оказывает существенное влияние на динамику различных
психофизиологических реакций и на вербальную оценку стимула.
1.
Брунер Дж. Психология познания. М.,
1977.— 412 с.
2.
Гласе Дж., Стэнли Дж. Статистические методы в педагогике и психологии.
М., 1976.— 495 с.
3.
Кочубей Б. И. Влияние генотипа и среды на параметры вызванных
потенциалов при ориентировочной и оборонительной реакциях // Вопр. психол. 1983. № 1. C. 141 —144.
4.
Кочубей Б. И. Индивидуальные особенности привыкания вызванных
потенциалов как отражение стратегии адаптации к стрессу // Психологические
проблемы индивидуальности. М., 1983. C. 36—37.
5.
Кочубей Б. И., Шеварев Ю. Н. Артефакты мигания
и их влияние на зависимость амплитуды вызванных потенциалов от интенсивности
стимула // Физиология человека. 1981. Т.
7. № 6. С. 1082—1086.
6.
Пучинская Л. М. Субъективность
восприятия и вызванный потенциал. М., 1978.— 149 с.
7.
Рутман Э. М. Функциональное значение
позднего положительного колебания вызванного потенциала // Физиология человека.
1980. Т. 6. № 4. C. 707—711.
8.
Соколов Е. Н. Восприятие и условный рефлекс. М., 1958.— 332 с.
9.
Соколов Е. Н. Ориентировочный рефлекс как информационный регулятор //
Ориентировочный рефлекс и проблемы рецепции в норме и патологии. М., 1963. C. 1—20.
10.
Стреляу Я. Роль темперамента в
психическом развитии. М., 1982.— 232 с.
11.
Черниговский В. Н., Мусящикова С. С., Синяя М. С, Мокрушин А. А. Привыкание в висцеральных системах. Л., 1980.— 242 с.
12. Begleiter
H., Porjesz В., Yerre С., Kissin В. Evoked potential correlates of expected stimulus intensity // Science.
1973. V. 179. P. 814—816.
13. Birchall
P. M. A., Claridge G. S. Augmenting-reducing of
the visual evoked potential as a function of changes in skin conductance level
// Psychophysiology. 1979. V. 16. N 5. P. 482—490.
14. Brown W. Conceptions of perceptual defence.
15. Buchsbaum
M. Self-regulation of stimulus intensity // Consciousness and
self-regulation. N.Y.: Plenum Press, 1976. N 1. P. 101 — 135.
16. Buchsbaum
M., Silverman J. Stimulus intensity control and the cortical evoked
response // Psychosom. Med. 1968. V. 30. N 1. P.
12—22.
17. Byrne D. Repression-sensitisation as a dimension of personality // Progress in
experimental personality research. N.Y.: Acad. Press, 1964. P. 169—220.
18. Callaway E. Habituation of averaged
evoked potentials in man // Peeke H.V.S., Herz M.J. (eds). Habituation.
N.Y.: Acad. Press, 1973. V. 2. P. 153—174.
19. Courohesne
E. Changes in P3 waves with event repetition // EEG Clin.
Neurophysiol. 1978. V. 45. N 6. P. 754—766.
20. Davis H., Osterhammel
R. A., Weir С. С., Giedigen
D. B. Slow vertex potentials: Interactions among
auditory, tactile, electric, and visual stimuli // EEG Clin.
Neurophysiol. 1972. V. 33. N 5. P. 537—545.
21. Graham F. К. Habituation and dishabituation
of responses innervated by the autonomic nervous system // Peeke
H.V.S., Herz M.J. (eds).
Habituation. N.Y.: Acad. Press, 1973. V. 1. P. 163—218.
22. Grzegolowska-Klarkowska
H. J. Use of defence mechanisms as determined by
reactivity and situational level of activation // Polish Psychol.
Bull. 1980. V. 11. N 3. P. 155—163.
23. Hilgard
E. Toward a neo-dissosiation theory // Perspect. Biol. Med. 1974. V. 17. N 3. P. 301—316.
24. Megela
A. L., Teyler T. J. Habituation of the human
evoked potentials // J. Compar. Physiol.
Psychol. 1979. V. 93. N 6. P. 1154—1170.
25. Näätänen
R., Michie P. T. Early selective attention
effects on the evoked potentials // Biol. Psychol.
1979. V. 8. N 1. P. 81 — 136.
26. O'Gorman J. G. Individual differences
in habituation of human physiological responses // Biol. Psychol.
1977. V. 5. N 4. P. 257—318.
27. Picton
T. W., Hillyard S. A., Galambos
R. Habituation and attention in the auditory system // Handbook of sensory
physiology. N.Y.: Acad. Press, 1976. V. 5. P. 344—381.
150
28. Raskin
D. С. Orienting and defensive reflexes and
conditioning // Black A.H., Prokasy W.F. (eds).
Classical conditioning. N.Y.: Acad. Press, 1972. P. 269—289.
29. Roth W. Т., Ford J. M., Kopell B. S. Long-latency evoked potentials and reaction time // Psychophysiology.
1978. V. 15. N 1. P. 17—23.
30. Routtenberg
A. The two-arousal hypothesis // Psychol. Rev.
1968. V 75. N 1. P. 51—80.
31. Thompson R. F., Spenser W. A. Habituation:
A model phenomena for the study of neuronal substrates of behavior // Psychol. Rev. 1966. V. 73. N 1. P. 16—43.
Поступила в редакцию 11.X 1984 г.