107
ПОПУЛЯЦИОННО-ГЕНЕТИЧЕСКИЙ АНАЛИЗ
НЕКОТОРЫХ ПАРАМЕТРОВ ЦВЕТОВОГО ВОСПРИЯТИЯ
К.Б. БУЛАЕВА, С.А. ИСАЙЧЕВ
Одним из важнейших аспектов проблемы соотношения биологического и социального в развитии человека является выявление взаимоотношений наследственности и среды в ходе онтогенетического развития как отдельных психических процессов, так и психики человека в целом. Влияние наследственности человека на его психическое развитие в значительной мере маскируется, а иногда полностью элиминируется влиянием социальных, экологических, половых и возрастных факторов. Кроме того, формируемые в процессе собственной деятельности индивида морфофункциональные мозговые структуры качественно отличаются от исходного материального субстрата, организация которого находится под значительным генетическим контролем. Все это делает задачу обнаружения влияний генотипа на психическое развитие человека чрезвычайно сложной.
В современной психогенетике основное внимание исследователей обращено прежде всего на выявление роли генетических факторов, обусловливающих формирование и развитие таких интегральных характеристик психики и высшей нервной деятельности, как интеллект, темперамент, восприятие, память, ЭЭГ, свойства нервной системы и т.д. Как правило, в исследованиях подобного типа констатируется лишь примерное соотношение генотипа и среды, оказывающих влияние на формирование изучаемых компонентов психической деятельности. Сопоставление средовых и генетических детерминант имеет важное значение для решения теоретико-психологических вопросов проблемы соотношения биологического и социального в развитии человека. Такая констатация влияния генотипа не дает возможности выделить из изучаемого параметра психической деятельности те звенья, которые можно с большей вероятностью назвать наследственно обусловленными, не позволяет выявить генетические механизмы нейрофизиологических процессов, являющихся материальным субстратом процессов психических.
Более перспективным может оказаться поиск и выбор в качестве объекта генетического анализа таких компонентов психической или высшей нервной деятельности, которые имеют явную генетическую детерминацию и влияют на формирование более сложных свойств и параметров, функционально связанных с этими компонентами. Кроме того, физиологические механизмы этих компонентов должны быть достаточно хорошо известны, так как действие генотипа сказывается именно на них. Такие признаки, физиологический механизм и генетическая природа которых хорошо изучены, носят в генетике название маркеров.
Одним из известных параметров цветового восприятия, удовлетворяющих этим условиям, является феномен аномальности цветового зрения — так называемая цветовая слепота. Выделяют три типа дефектов цветового зрения: протанопию, дейтеранопию, тританопию и соответствующие им менее тяжелые нарушения — протаномали-протаномалию, дейтераномалию и тританомалию. Почти окончательно установлено, что протанопический и дейтеранопический дефекты возникают вследствие мутаций по двум различным, но весьма тесно сцепленным локусам Х-хромосомы. Родословные лиц с аномалиями или дефектами цветового зрения показывают, что эти нарушения наследуются как рецессивный, сцепленный с полом признак. Нормальное цветовосприятие является доминантным по отношению к аномальному. Легкость определения и сравнительно частая встречаемость аномалий (от 2 до 8 % в Европе [3; 35]) позволяют использовать их в качестве генетического маркера при описании генетических структур популяций.
По вопросу о наследуемости нормального цветового восприятия систематического изучения не проводилось, существуют лишь отдельные данные о наследственных особенностях некоторых параметров цветового восприятия. Так, ряд исследователей [2], [6] показали, что среди людей, обладающих нормальным цветовым зрением, можно выделить две группы, различающиеся между собой тем, что субъективному ощущению чистого зеленого цвета у них соответствуют две различные по длине волны точки спектра. Первая группа имеет среднюю арифметическую этого признака в области спектра с длиной волны около 516 нанометров1, вторая — с длиной волны около 526 нм. Позднее
108
высказывалась гипотеза [4] о наследуемости этого свойства нормального цветового зрения. Для ее подтверждения были приведены экспериментальные данные.
Таким образом, можно полагать, что некоторые параметры цветового восприятия имеют генетическую детерминацию и они, в известной мере, могут влиять на формирование нормального восприятия цвета.
Основная цель настоящей работы состояла в исследовании ряда параметров цветового восприятия и выявлении из них наиболее элементарных, генетически обусловленных, таких, которые можно будет использовать в последующих исследованиях в качестве генетических маркеров. Выявление таких признаков и применение к ним методов генетического анализа позволит определить их тип наследования, локализацию и взаимосвязи всех моногенных и полигенных систем, оказывающих влияние на формирование и развитие цветового восприятия человека.
Известно, что генотипическая архитектоника полигенных и моногенных систем, лежащих в основе формирования структур и функций организма, зависит от генетической структуры популяции в большей степени, чем от генетической природы самого признака. Поэтому генетический анализ любого количественного признака с полигенной системой детерминации требует прежде всего учета генетической структуры популяции в целом. Руководствуясь этими соображениями в качестве основного объекта исследования, были выбраны малые популяции человека — так называемые изоляты. Изолятами называют как отдельные малые, изолированные в силу каких-либо причин популяции, так и небольшие группы, входящие в состав популяции, при условии, что в отношении размножения соответствующие группы остаются какой-то отдельностью.
Именно сохранение из поколения в поколение постоянства круга вступающих между собой в брак и обусловливает генетические различия между популяциями. Уникальная генетическая структура каждого отдельного изолята позволяет выявить взаимоотношения генетических и средовых факторов, влияющих на изменчивость и наследуемость признака в зависимости от различий в генетических структурах, так как частота систем аллельных генов, особенности их взаимодействия в конкретных популяциях отражаются на фенотипическом проявлении изучаемого признака. Кроме того, популяционный подход позволяет выявить весь диапазон вариабельности признака, что дает возможность перейти от относительной оценки параметров признака в пределах одной экспериментальной выборки к оценке абсолютной [1].
Для экспериментального исследования нами были выбраны относительно самостоятельные и на первый взгляд не связанные между собой феномены цветового восприятия: дифференциальная чувствительность в синей, зеленой и красной областях спектра, подравнивание желтого цвета к смеси красного и зеленого (уравнение Релея), аномалии цветового зрения, определение точек спектра, соответствующих субъективному образу «чистого зеленого» и «чистого желтого» цветов.
Таблица 1
ЧАСТОТА АНОМАЛИЙ ЦВЕТОВОГО ЗРЕНИЯ, ЧАСТОТЫ ГЕНОВ σ1 И σ2,
ОБЩЕПОПУЛЯЦИОННЫЕ СРЕДНИЕ ВЕЛИЧИНЫ— М (В НАНОМЕТРАХ) ПРИЗНАКА «ВОСПРИЯТИЕ ЧИСТОГО ЗЕЛЕНОГО ЦВЕТА»
В РАЗЛИЧНЫХ ПОПУЛЯЦИЯХ ДАГЕСТАНА
|
Популяция |
Размер выборки |
Частота аномалий, % |
G1 |
G2 |
M |
|
|
у мужчин |
у женщин |
|||||
|
Тинди |
132 |
4,17 |
— |
0,711 |
0,289 |
519,4 |
|
Н. Мехельта |
101 |
21,43 |
3,39 |
0,667 |
0,333 |
520,6 |
|
Чуртах |
104 |
— |
— |
0,641 |
0,359 |
523,4 |
|
Ботлих |
158 |
10,67 |
3,61 |
0,478 |
0,522 |
523,4 |
|
Экзогамная группа |
169 |
9,52 |
2,35 |
0,530 |
0,470 |
522,0 |
МЕТОДИКА
Объектом исследования были выбраны изолированные популяции Дагестана, характеризующиеся высоким коэффициентом инбридинга (характеризует родственные браки разной степени) и эндогамией. Межпопуляционные различия в генетических структурах исследованных популяций определялись по совокупности генетических маркеров. Исследование проводилось на контингенте учащихся VIII—X классов и их родственниках. Все испытуемые проходили предварительное исследование, целью которого являлось выявление лиц с дефектами цветового зрения. Диагноз аномалий ставился с помощью полихроматических таблиц Рабкина [2] и аномалоскопии на аномалоскопе Раутиана АН-59. Данные, полученные от испытуемых с выявленными дефектами цветового зрения, обрабатывались отдельно и не входили в общепопуляционные вариационные распределения признаков.
Оценка дифференциального порога в красной, зеленой и синей областях спектра, а также составление уравнения Релея (подравнивание желтого цвета спектра (589 нм) к смеси спектральных красного (670 нм) и зеленого (535 нм) проводились на аномалоскопе Раутиана АН-59. Определение точек спектра, соответствующих субъективному образу «чистого зеленого» и «чистого желтого» цветов, проводилось на спектральном аномалоскопе Рабкина, позволяющем испытуемому воспринимать все разнообразие тонов зеленого и желтого цветов спектра в диапазонах 495—550 нм (для зеленого) и 560—600 нм (для желтого). После каждой установки выбранного испытуемым цвета влияние хроматической адаптации снималось путем адаптации к белому цвету.
Были обследованы четыре изолированные популяции и экзогамная выборка, в которую входили дети от смешанных (межаульных) браков. Общее количество обследованных — 664 человека.
109
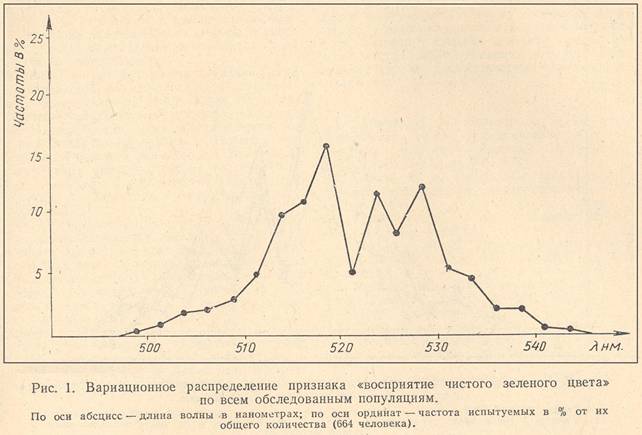
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В табл. 1 представлены данные о частоте аномалий цветового восприятия в обследованных изолятах. Частота дефектов заметно выше среди мужчин, чем среди женщин, что и следовало ожидать от сцепленного с полом рецессивного признака. Показаны основные типы выявленных аномалий дейтеранопического и протанопического характера. Монохроматизм и тританопические дефекты в обследованных изолятах не обнаружены. Из 35 обнаруженных лиц с аномалиями 20 человек дейтанов и 15 протанов. Частота дефектов цветового зрения сильно варьирует в различных популяциях — от 0 до 21 % пораженных в группе мужчин и от 0 до 3,6 % в группе женщин. Эти данные значительно отличаются от литературных о распространенности таких дефектов, согласно которым в Европе процент пораженных среди мужчин составляет 7—8 % [3].
Такие резкие различия в концентрации рецессивного аллеля красно-зеленой слепоты можно объяснить механизмами изоляции, эффектом родоначальника и спецификой протекания генетико-автоматических процессов, которые могут глубоко дифференцировать генный состав малых популяций и вызвать в их системе серьезные генетические различия.
Для выявления характеристик распределений изучаемых признаков проводился их статистический анализ. С этой целью строились вариационные кривые по каждому признаку, находились основные статистики распределений и проводилось сопоставление вариационной кривой каждого признака с кривой нормального распределения (аппроксимация) по критерию χ2. За исключением признака «восприятие чистого зеленого цвета», вариационные кривые всех изучаемых признаков не отличаются от кривой нормального распределения, и эти признаки могут быть отнесены к разряду количественных, т.е. таких, в которых межиндивидуальные различия выражаются прежде всего степенью проявления признака, которую можно измерить, т. е. приписать, определенное количественное значение.
Вариационное распределение признака «восприятие чистого зеленого цвета» значительно отличается от кривой нормального типа и имеет бимодальный характер, т. е. выделяются две несколько перекрывающиеся (трансгрессирующиеся) группы, в каждой из которых значение признака проявляется по своему (рис. 1). Среднее значение точки спектра, соответствующее субъективному восприятию «чистого» зеленого цвета, в первой группе равно 5J5 нм, во второй — 529,4 км. Такая бимодальность проявляется в распределении данного признака в каждой из обследованных популяций (рис.2). Между популяциями наблюдаются различия в средних значениях и дисперсиях.
Бимодальный характер вариационной кривой признака «восприятие чистого зеленого цвета» может быть вызван как генетическими механизмами, так и наличием полового или возрастного диморфизма, связанного с причинами генетической и средовой природы. Для выявления полового и возрастного диморфизма проводилось сравнение выборок взрослых и детей, а также мужчин и женщин в обследованных изолятах, по критерию Стьюдента и Фишера. Систематических различий между детьми и взрослыми, мужчинами и женщинами не обнаружено. Визуальный анализ вариационного распределения восприятия «чистого» зеленого цвета
110
позволяет выделить в выборке женщин промежуточную группу со средней длиной волны около 521—523 нм. Таким образом, в каждом изоляте можно выделить два фенотипа мужчин G1 и G2 и три фенотипа женщин G11 G12, G22. Полученные данные вполне согласуются с гипотезой Валера (1967) о том, что бимодальный характер вариационной кривой восприятия «чистого» зеленого цвета определяется наследственными факторами, которые имеют один локус в одном цистроне Х-хромосомы с двумя мутантными кодоминантными генами. С целью проверки данного предположения нами был проведен анализ сегрегации этого признака в семьях. Результаты такого сегрегационного анализа в различных типах семей приводятся в табл. 2 и показывают хорошее согласование с указанным выше предположением. Так, из 122 пар «мать—сын» только две пары, а из 68 пар дочерей и двух родителей только одна пара не согласуются с вышеуказанной гипотезой, что, по-видимому, является артефактом.
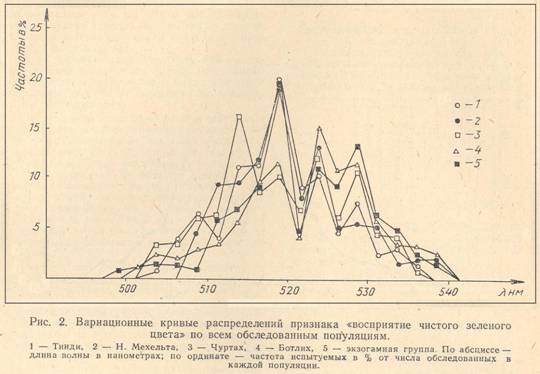
Различия между популяциями выражаются спектральным сдвигом среднего значения признака и соотношением групп испытуемых с разными фенотипами: G1 и G2 — у мужчин, G11 G12, G22 — у женщин. На основе частот фенотипов мужчин и женщин вычислялись частоты генов G1 и G2 в обследованных изолятах и об-щепопуляционные средние значения рассматриваемого признака в них (см. табл. 1). Из представленных в табл. 1 данных можно заметить, что частоты генов G1 и G2 значительно варьируют в разных изолятах. Проверка однородности популяций по критерию χ2 показала, что все четыре изолята достоверно различаются по частотам генов G1 и G2, т.е. генетическую неоднородность изучаемых популяций по содержанию данных генов.
Таблица 2
АНАЛИЗ СЕГРЕГАЦИИ ПРИЗНАКА «ВОСПРИЯТИЕ ЧИСТОГО ЗЕЛЕНОГО ЦВЕТА»
В РАЗЛИЧНЫХ ТИПАХ СЕМЕЙ
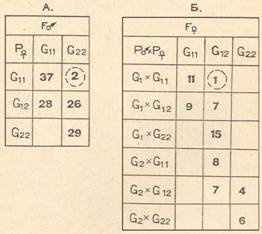
Примечания. A — G11 G12, G22 —фенотипы матерей; G11, G22— фенотипы сыновей,
Б —Р×Р — фенотипы родителей; G11 G12, G22 — фенотипы дочерей.
ВЫВОДЫ
1. Исследуемые признаки (параметры) цветового восприятия человека могут быть разделены
111
на количественные с непрерывным и качественные с дискретным популяционным распределением; по отношению к последним возможно применение менделеевского анализа и определение типа наследования и локализации их генетических детерминант.
2. Популяционно-генетический анализ ряда параметров цветового восприятия человека показал, что особенности генетической структуры конкретной популяции могут отражаться как на фенотипической, так и на генотипической изменчивости изучаемых признаков. Поэтому при определении влияния генетического фактора на формирование признака (параметра) необходимо учитывать особенности генетической структуры популяции, в которой проводится исследование.
3. Частота аномалий цветового зрения в некоторых обследованных изолятах значительно отличается от средней частоты аномалий в Европе. Такая высокая концентрация рецессивного гена красно-зеленой слепоты свидетельствует о специфике и интенсивности протекания микроэволюционных процессов в генофондах малых, изолированных популяций человека и их глубокой дифференциации.
4. Дискретный характер вариационного распределения признака «восприятие чистого зеленого цвета» обусловлен влиянием генетического фактора — двумя кодоминантными генами G1 и G2, имеющими один локус в Х-хромосоме. Такой характер распределения признака обнаружен в различных по своему генофонду популяциях. Межпопуляционные различия выражаются смещением средних значений популяционных распределений признака, что объясняется генетической неоднородностью популяций по содержанию генов G1 и G2.
1. Дубинин Н. П., Булаева К. Б. Изучение генетических основ индивидуальности в популяциях человека. — Доклады АН СССР. 1982. Т. 265. № 2. С. 470—473.
2. Рабкин Е. Б. Полихроматические таблицы для исследования цветоощущения. — М., 1971. —244 с.
3. Харрисон Дж. и др. Биология человека. — М., 1979. — 609 с.
4. Waaler G. H. M. Heredity of two normal types of color vision. Nature. 1967. V. 215. P. 688.
5. Richards W. Differences among color normals: Classes I and II // J. Opt. Soc Am. 1967. V. 57. N 08. P. 1047—1055.
6. Rubin M. L. Spectral hue loci of normal and anomal trichromates // Am. J. Ophthalmol. 1961. V. 52. P. 166—172.
Поступила в редакцию 19.IV 1983 г.