130
НАУЧНЫЕ ОБЗОРЫ
ВИДИМОЕ ДВИЖЕНИЕ: ФЕНОМЕНОЛОГИЯ, ОСНОВНЫЕ
ДЕТЕРМИНАНТЫ, МЕХАНИЗМЫ1
Ю. Н. МИРТОВ
Видимое, или
кажущееся, движение (его также называют стробоскопическим) — зрительная иллюзия
движения — представляет собой классический объект исследований в психологии
зрительного восприятия. Считается, что общая теория зрительного восприятия
движения должна включать в себя объяснение видимого движения. С другой стороны,
изучение видимого движения может помочь в построении такой теории [5], [45].
Термином «видимое движение» охватывается целый класс феноменов,
возникающих при некотором режиме предъявления неподвижных паттернов а, d, с, ..., n, имеющих пространственную диспаратность. Каждый отдельный паттерн может состоять из
одного или нескольких элементов или фигур. В дальнейшем такая стимульная
ситуация будет называться стробоскопическим стимулом или просто стимулом. Режим
предъявления заключается в кратковременной поочередной экспозиции паттернов или
в постепенном изменении их яркости. Переход от одного типа видимого движения к
другому обусловлен параметрами как стробоскопического стимула, так и режима
предъявления. От этих же параметров (детерминантов) зависит в первую очередь и
качество того или иного типа видимого движения (степень близости к впечатлению,
вызываемому действительным перемещением паттерна — реальным движением).
Перечислим основные детерминанты типа и качества видимого движения:
1. Время экспозиции паттернов (D). Исчисляется, как правило, десятками миллисекунд
(мс), но может достигать и нескольких секунд.
2. Временной интервал между экспозициями паттернов (мс). Исследования
показали целесообразность различения двух временных интервалов. Межстимульный интервал (t) — время между концом экспозиции предыдущего
паттерна и началом экспозиции последующего. Асинхронность предъявления
паттернов (Т) — время между началом экспозиции предыдущего паттерна и началом
экспозиции последующего. Следовательно, Da + t=T и если Da>T,
то t<0.
3. Яркость паттернов (I).
В том случае, когда фон, на котором предъявляется стробоскопический стимул,
также имеет некоторую яркость, более важным детерминантом оказывается абсолютное
значение контраста фигура — фон.
4. Межстимульное расстояние — расстояние
между паттернами. Обычно измеряется зрительным углом между центрами паттернов (α). Когда лишь некоторые элементы из каждого паттерна
участвуют в движении (образуют стробоскопические пары), более важным
оказывается межстимульное расстояние в
стробоскопических парах.
5. Размер паттернов (или фигур, составляющих стробоскопические пары).
Также обычно измеряется зрительным углом, в котором заключен паттерн (β). Причем имеет значение только размер
паттерна по его сечению траекторией движения.
На видимое движение оказывают влияние и другие характеристики
стробоскопического стимула: форма паттернов и отличие паттернов по форме или
ориентации, видимое межстимульное расстояние и
видимый размер паттернов или фигур, структурированность фона, предметность
паттернов и т. п., и характеристики, относящиеся к наблюдателю: возраст, пол,
жизненный опыт, тренированность, распределение внимания, в конечном счете —
установка. Кроме того, могут иметь вес такие факторы, как стимуляция со стороны
других модальностей или дополнительные задачи, выполняемые наблюдателем. Также
большую роль играют условия предъявления стробоскопического стимула: бинокулярно, монокулярно или дихоптически,
локализация стимула на сетчатке. Некоторые из этих факторов будут обсуждены
ниже. В основном же обзор будет ограничен ситуациями, в которых в качестве
паттернов применяются простые геометрические фигуры на неструктурированном
фоне, предъявляемые в парафовеальной области
сетчатки. В этом случае главными факторами, определяющими тип и качество
видимого движения, являются перечисленные выше пять параметров
стробоскопической стимуляции и установка наблюдателя.
ТИПЫ СТРОБОСКОПИЧЕСКОГО СТИМУЛА И ФЕНОМЕНОЛОГИЯ
Для упрощения изложения «стандартизируем» стробоскопический стимул.
Пусть паттерны предъявляются на неосвещенном экране, расположенном во
фронтальной плоскости и удаленном от глаза на расстояние 1 м. Пусть яркость
фигур имеет некоторое среднее значение, α =2°, а
β =0,5°. Теперь важными детерминантами остаются
только время экспозиции паттернов и t
или Т. Если значения параметров стимуляции будут существенно отличаться от
установленных, то они будут указаны. В дальнейшем будут употребляться некоторые
условные обозначения. Запись «За, b»
расшифровывается так: стимул, состоящий из двух паттернов а и b, паттерн а предъявляется
131
первым и
содержит три фигуры, паттерн b
вторым и содержит одну фигуру. Запись a1 — b3» означает: движение по траектории от фигуры № 1 паттерна а до фигуры №
3 паттерна b.
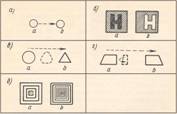
Рис. 1. Двухфигурные
стробоскопические стимулы. Латинскими буквами указан порядок предъявления
фигур, стрелками — направление воспринимаемого движения.
а — простейший стимул; б — стимул для демонстрации реверсивного
видимого движения (Anstis, 1970); в — стимул для демонстрации пластичного изменения формы (Kolers Pomerantz, 1971); г — стимул для демонстрации поворота ригидной
фигуры в пространстве (Kolers, Pomerantz, 1971); д — стимул для
демонстрации движения в глубину (Bernstein et al., 1971).
Основы типологии видимого движения были сделаны Вертхаймером
в его фундаментальном в этой области исследовании [66]. Основной переменной, от
которой зависит тип видимого движения, он считал t. В диапазоне изменения t от 31 мс до 322 мс (Da, b = 7 ÷ 33 мс) он выделил следующие
качественно отличные восприятия, сопровождающие предъявление двухфигурного стробоскопического стимула a, b (рис. 1, а):
1. Симультанное восприятие фигур а
и b: одновременно воспринимаются две неподвижные
фигуры ( t ≈30 мс).
2. Частичное движение.
а) Внутреннее движение (intramembral) — независимое мелькание обеих фигур. Внутри
одной или обеих фигур наблюдается дрожание контура или флуктуации яркости (t несколько больше, чем 30 мс).
б) Двойное движение (bimembral) — фигура проходит только часть траектории.
В середине траектории существует больший или меньший разрыв (t ≈ < 45 мс, а также t ≈ > 60 мс).
в) Одиночное движение (unimembral) — только одна из двух фигур движется,
другая или неподвижна, или не воспринимается вовсе (30 мс ≈ < t ≈ < 200 мс).
3. Оптимальное движение.
Равномерное движение фигуры от места первого предъявления (а) к месту второго (b), причем фигура последовательно видна во
всех промежуточных положениях, а ее яркость остается неизменной (t ≈ 60 мс). Кенкел [44] назвал этот тип
движения «бета-движением».
4. Фи-движение
Восприятие перемещения одной фигуры из положения а в положение b отсутствует, но сохраняется некоторое
впечатление движения, перехода. Это чистое, безобъектное движение —
туннель-эффект (во всем диапазоне изменения t, но наиболее вероятно при t, немного больших 30 мс и немного меньших 200
мс). Иногда видимое движение в целом называют фи-феноменом
и даже фи-движением. Фи-движение
может по-разному интерпретироваться наблюдателями. Так, другими авторами оно
было описано как омега-движение при периодической последовательности вспышек
фигур а и b (Da, b=200 мс, t = 0) наблюдателю кажется, что фигуры по
очереди закрываются некоторым непрозрачным движущимся объектом, т. е. движение
приписывается самому «туннелю». Переход от восприятия бета-движения
к омега-движению зависит от колебаний установки наблюдателя
[59], [63], [69].
5. Сукцессивное восприятие обеих фигур.2 Восприятие, соответствующее
действительной пространственно-временной организации стимуляции (t ≈ 200 мс).
Вертхаймер отмечал, что видимое движение очень
лабильный феномен и что то или иное значение t определяет большую или меньшую вероятность
того или иного восприятия. Кроме того, он считал, что только оптимальное
движение можно отождествлять с реальным движением. Все остальные типы
восприятия — это не типы, не стадии оптимального движения, это движутся другие
объекты.
В связи с этим возникает проблема измерения выраженности или качества
видимого движения. Допустим, нас интересует качество видимого движения как
функция от t. Можно ли считать, что
при t = 30 мс оно ниже, чем
при t = 60 мс? Согласно Вертхаймеру, правильнее говорить о степени благоприятности
значения t для восприятия бета-движения. Величина вероятности восприятия бета-движения как функция от t для кратких Da ≈ < 1000 мс описывается U-образной кривой, достигающей максимума
обычно при значениях Т ≈ 100 мс, а для долгих Da ≈ > 100 мс при t ≈ 0 [6], [34], [46]. Но можно отнестись к восприятиям типа l ÷ 5 как к качественным стадиям бета-движения. Никто не отстаивал эту точку зрения и не
ставил под сомнение позицию Вертхаймера, тем не менее
шкалирование качества видимого движения утвердилось в
практике исследований. Обычно для этой цели применяют метод рангового
упорядочивания [2], [23], [42], [61], нами был апробирован метод парных
сравнений [8]. Другая проблема измерения качества видимого движения — проблема
содержания и уровня критерия наблюдателя. Между двумя наблюдателями может не
быть согласия по вопросу о качестве движения. Более того, критерий наблюдателя
может изменяться вместе с изменением значений параметров стимуляции [43], [45].
Какое же движение следует считать качественным? Десильва,
который поставил этот вопрос, дал на него радикальный ответ [31]. Он считал,
что качественное видимое движение это такое впечатление движения, которое
действительно неотличимо от реального движения. Поэтому метод Десильвы — прямое сравнение видимого движения и реального
движения при всех прочих равных условиях. «При прочих равных условиях» означает,
132
что форма,
яркость, размеры реально движущейся фигуры идентичны фигурам а и b. Кроме того, равны межстимульное
расстояние в стробоскопическом стимуле и амплитуда реального движения. Теперь
остается придать реальному движению такую же скорость, какую имеет видимое
движение. Но как раз определение скорости видимого движения и представляет собой
проблему. На первый взгляд, V = α / t, поскольку в течение Da
фигура «стоит» на одном месте. Однако видно, что она действительно стоит,
только если Da ≈
> 100 мс, а если Da ≈ < 100 мс, то фигура воспринимается сразу в движении [42],
[43], [46]. Поэтому если Da ≈ < 100 мс, то V
= α / T [31], [45], а если Da ≈ > 100 мс, то V
= α / (t + x),
где x — некоторая неизвестная
величина. То, что эта величина существует, доказывает тот факт, что
бета-движение воспринимается в этих условиях, даже если t ≤ 0 [37], [51], [66]. Для определения
величины х
следует выяснить временные характеристики процесса восприятия видимого
движения.
Колебательное бета-движение. Если наблюдателю предъявляется
периодическая последовательность вспышек а и b так, что ta-b = tb-a, то он видит возвратно-поступательное
движение фигуры. Если у испытуемого при этом вызывают каким-либо способом
ассоциации с раскачивающимся маятником, то он видит маятникообразное
бета-движение — дугообразное искривление траектории движения в вертикальной или
в горизонтальной плоскости [36], т. е. в последнем случае траектория движения
«выходит в пространство» из плоскости дисплея. Существует также комбинация из
прямого и дугообразного видимого движения. Если ta-b = 50 мс, а tb-а = 150 мс, Da,b = 50 мс, то наблюдатель в направлении а—b видит прямое бета-движение, а в направлении b—а — дугообразное движение в горизонтальной
плоскости [6]. При продолжительном наблюдении колебательного бета-движения становятся заметными его флуктуации:
спонтанные переходы от одного типа восприятия к другому во всем диапазоне от симультанности до сукцессивности.
Причем общее время восприятия бета-движения в течение
определенного промежутка времени является функцией от значений параметров
стимуляции [6], [48]. Тренированный наблюдатель может также произвольно
настраиваться на тот или иной тип восприятия [51].
К маятникообразному видимому движению можно
отнести также феномен «блуждающего» (Wandering) света, или W-феномен
[40]. Обе фигуры циклически (с частотой от 0,2 до 3 Гц) и плавно изменяют в противофазе
свою яркость. Наблюдатель видит маятникообразное
движение фигуры в горизонтальной плоскости, причем ее видимая яркость
сохраняется постоянной.
Реверсивное движение. Если яркость фигуры b значительно превышает яркость фигуры а, то
при последовательности вспышек a,
b наблюдатель видит движение
фигуры в направлении b—а.
Это так называемое дельта-движение [47]. Другой тип реверсивного движения описан
Колерсом [45]. Если Dа = 250 мс, Db = 10 мс, t =10
мс, а ≈ < 15', Iа,
b = 0, фона I = 4 млк, то при предъявлении
a, b воспринимается движение по траектории а—b—а. Еще один тип реверсивного движения
получен Энстисом [12]. Iа постепенно уменьшается, а Ib синхронно увеличивается (αa—b = 0). Наблюдатель видит движение буквы «Н»
влево, хотя Нb
смещена относительно На вправо на 15' (рис. 1 б).
Движение изменяющейся формы и
движение ригидной формы.
Если фигуры а и b имеют
различную форму, то при бета-движении наблюдается
пластичная деформация фигуры а, превращение ее в фигуру b по мере движения из одного положения в
другое (рис. 1 в) [51], [53], [66]. Если фигуры а и b отличаются по своей ориентации в
пространстве (рис. 1, г), то при t
= 50 мс наиболее вероятно восприятие движения изменяющейся формы, а при t = 260 мс [46] — восприятие движения ригидной
формы: вращение фигуры в плоскости дисплея или в пространстве в процессе
движения (Da,b = 25÷400 мс) [3], [65].
Гамма-движение. Стимул для демонстрации гамма-движения
состоит только из одной фигуры. Гамма-движением
называется впечатление расширения — сжатия при быстром увеличении — уменьшении
яркости фигуры или просто при ее включении — выключении [44].
Движение в глубину. Изучая гамма-движение, получаемое при
увеличении — уменьшении яркости фигуры, Ньюман
обнаружил, что его можно рассматривать и как движение вперед — назад от
наблюдателя [52]. Бартли и Миллер получили движение в
глубину при следующих условиях: α = 0, D a, b = 5 мс, Т = 10 мс, Ia >
Ib. Это движение было названо ими «adab» [17]. Кроме того, движение в глубину получают,
предъявляя на одном месте (α = 0) различные по
размеру фигуры. В зависимости от установки наблюдателя в этих условиях может
восприниматься и движение расширения — сжатия [52], [67].
Альфа-движение — впечатление расширения— сжатия или
движения в глубину при предъявлении фигур, имеющих не действительное, а видимое
отличие в размерах, например фигур Мюллера — Лайера
[44], или отличие по видимой глубине, как, например, в работе Бернштейн и др.
(рис. 1, д) [18]. Этими же авторами описан способ
оценки выраженности видимого движения по направлению к наблюдателю,
заключающийся в измерении силы ориентировочного рефлекса.
II. Трехфигурные
стимулы.
а) Стимулы а, 2b и 2а,
b.
Эффект расщепления фигуры а. Стимул для демонстрации этого эффекта был
разработан Вертхаймером [66] (рис. 2, а). Однако он
считал движение в этих условиях невозможным и рассматривал его как один из
вариантов фи-движения. Ряд других исследователей,
напротив, считают, что такое движение хорошо наблюдается [8], [29], [45], [51],
[61]. Более наглядный стимул для демонстрации эффекта расщепления (его также
называют билатеральным - движением) представил Нейгауз, получив движение и в обратном
133
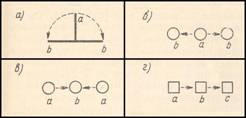
Рис.
2. Трехфигурные стробоскопические стимулы. Обозначения
те же, что на рис. 1.
а — стимул для
демонстрации эффекта расщепления (Wertheimer,
1912); б, в — стимулы для демонстрации билатерального движения и движения,
обратного билатеральному (Neuhaus, 1930); г — стимул для демонстрации тау-эффекта (Geldreich, 1934).
направлении в
стимуле 2 a, b (рис. 2, b и 2,
в) [51].
б) Стимул а, b, с
(рис. 2, г).
При предъявлении этого стимула воспринимается бета-движение а—b—с. Но его качество хуже, чем при
предъявлении стимула а, b
наблюдатель замечает дискретные фазы движения [45].
Тау-эффект. Если αa-b = αb-cс, a ta-b > tb-c, видимое αa-b больше видимого αb-с [33].
Каппа-эффект — эффект, обратный тау-эффекту.
Если ta-b = tb-c, а αа-b >αb-с то кажется, что ta-b > tb-c [28].
III. Многофигурные стимулы.
Многофигурные стимулы представляют собой различные комбинации из уже
описанных стимулов. Например, эффект расщепления фигуры может быть
продемонстрирован и в том случае, если паттерн b состоит из большего числа фигур, чем две
(рис. 3, а и б) [45]. Доказывая, что движения глаз не имеют отношения к
видимому движению, Вертхаймер разработал стимул 2а, 2b (рис. 3, ж). Наблюдатель видит одновременное
движение в противоположных направлениях: a1—b1 и а2—b2. Группировка фигур в стробоскопические пары зависит от установки
наблюдателя, которая может облегчаться или затрудняться значениями параметров
стимуляции. В приведенном примере фигуры группируются по принципу сходства
размеров. Другими факторами, определяющими группировку фигур и траекторию
движения, могут быть сходство по яркости, форме, ориентации, цвету,
пространственная близость фигур, принцип «хорошей» формы траектории [47].
Приведем пример группировки по цвету (рис. 3, з):
паттерн 2а (a1 — красная, а2 — зеленая)
совпадает в пространстве с паттерном 2b
(b1 — зеленая, b2 — красная). Наблюдатель видит движение красного круга (a1—b2) вправо и зеленого (а2—b1) влево [68].
Вращательное движение. При последовательности вспышек а, b, с, d, а (рис. 3, в) возникает восприятие одной
фигуры, движущейся по круговой траектории (принцип «хорошей» траектории) [26].
Вращательное движение получают также, предъявляя стимулы, показанные на рис. 3,
г, 3, д и 2, а. Направление движения при предъявлении
стимулов, изображенных на рис. 2, а и 3 г, зависит от установки наблюдателя [29],
[58], [66].
Эффект индукции. При чередовании паттернов 3а и 2b (рис. 3, е) центральная фигура паттерна а, у
которой нет пары в паттерне b,
также совершает колебательное движение, хотя выраженность этого движения
значительно меньше, чем в двух других парах [45].
Движение группы и движение
элемента. Паттерн 3а
сменяется паттерном 3b, причем фигуры а2 и а3 совпадают соответственно
с фигурами b1 и b2 (рис. 3, и).
Если t = 10 мс, то наблюдатель
видит движение одного элемента a1—b3 и два других — как неподвижные (Da,b = 200 мс; αa-b = 1˚, а если t = 80 мс, движение группы 3а — 3b [55]. Движение группы также называют эффектом
Тернуса [62].
Основные детерминанты бета-движения.
Механизмы видимого движения
Можно легко заметить, что почти все типы видимого движения являются
лишь более сложными вариантами бета-движения. В силу
своего фундаментального положения бета-движение представляет особый интерес.
Было бы чрезвычайно заманчиво выяснить влияние временных, пространственных и
энергетических характеристик стробоскопической стимуляции на бета-движение и их
взаимосвязь. Проблема детерминантов бета-движения
всегда связывалась с проблемой его механизмов. На этот счет существуют сотни
работ, однако известно все еще мало. Решение проблемы детерминантов бета-движения осложняется его большой зависимостью от установки
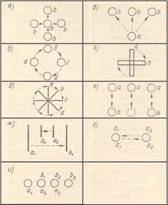
Рис.
3. Многофигурные стробоскопические стимулы. Обозначения те же, что на рис. 1.
а, б — стимулы для
демонстрации эффекта расщепления (Kolers, 1972); в, г, д — стимулы
для демонстрации вращательного движения (Brown, Voth, 1937; Robins, Shepard, 1977); e —
стимул для демонстрации эффекта индукции (Kolers, 1972); ж, з — стимулы для
демонстрации группировки фигур в стробоскопические пары: ж — по принципу
сходства размеров (Wertheimer, 1912); з — по принципу сходства цвета (a1 и b2—
красного цвета, а2 и b1 — зеленого) (Zapparoli, 1967); и — стимул для демонстрации движения группы
3а—3b и движения элемента a1—b3 (Pantle, Picciano, 1976).
134
наблюдателя.
Поэтому оказывается трудным, а порой и невозможным сопоставить результаты
различных исследований. Ниже мы попытались обобщить результаты, которые в
большей или меньшей степени выдержали последующую проверку. Не имея возможности
в рамках настоящей статьи сделать обзор всех точек зрения на механизмы видимого
движения [13], [45], мы кратко осветим лишь некоторые из них, имеющие связь с
нашей собственной концепцией.
В силу того что видимое движение — феномен, зависящий от многих
факторов, величина диапазона изменения какой-либо переменной, в котором
наблюдается бета-движение, зависит от значений других детерминантов. Диапазоны
же, в которых изменяются значения детерминантов, ограничены только разрешающей
способностью зрительной системы. Исключение составляют лишь t или Т и α, по
которым бета-движение имеет свои собственные пороги. Ниже будут указаны
предельные значения параметров стробоскопического стимула, известные нам по
литературным источникам.
Размеры фигур могут изменяться в широком диапазоне: от 3'
[19], а в случайно-точечных паттернах и меньше — 1,75' [49], до 20° и больше
[30]. Колерc считает, что существует
какой-то оптимальный β, но какой он и как
изменяется в связи с ретинальной локализацией и
другими детерминантами, никто не пытался установить [45]. Если α = const, то увеличение β позволяет расширить
диапазон изменения других переменных, в котором сохраняется восприятие бета-движения [38].
Яркость фигур может изменяться от пороговых значений [39]
до максимальных величин. Более важным детерминантом является значение разности Iа — Ib. То или иное ее значение может иметь самые
различные последствия. Значительно более яркая фигура может оказать маскирующее
влияние на другую, так что та вообще не будет воспринята [11]. При несколько
меньшей разнице, если Iа
< Ib возникает
дельта-движение [47]. С другой стороны, при средних значениях яркости Iа > Ib оказывает некоторое ухудшающее влияние на
бета-движение [48]. Студент Коффки Корте [47]
установил ряд попарных взаимоотношений между
детерминантами бета-движения, которые Коффка назвал законами. Корте объединил Da и Iа в одну общую переменную — интенсивность. Позже законы Корте были
скорректированы и здесь приводятся в этой более поздней редакции [5]. Первый
закон определяет связь между Iа и α: для сохранения бета-движения
при увеличении одного следует увеличить другое, т. е. Iа и α связаны между собой прямо
пропорционально. Второй закон устанавливает обратно пропорциональные отношения
между t и Iа Вопрос о степени корректности этих и других законов Корте остается
открытым. Многочисленные «ревизии» законов Корте [37], [50], [51], [64] не
привели к более точным формулировкам. По-видимому, правильным будет считать,
что они описывают взаимоотношения между параметрами стробоскопической
стимуляции в самом общем виде [45].
Время экспозиции фигур, межстимульный интервал и асинхронность включения фигур.
Время экспозиции фигуры b
не оказывает влияния на бета-движение [47], [51]. Db определяет только, как долго будет видна
вторая фигура и также ее яркость в пределах действия временной суммации. Время
экспозиции фигуры а может изменяться в широких пределах от 1 мс [19] до 10 с
[57], причем это отнюдь не предел. При увеличении Da восприятие бета-движения
становится более устойчивым [6]. Как указывалось выше, если Da ≈ < 100 мс, то
фигура воспринимается сразу в движении, а если Da ≈ > 100 мс, то фигура
сначала видна как неподвижная и уже затем начинает свое движение. Четвертый
закон Корте устанавливает обратно пропорциональные отношения между t и Da, т. е. фактически утверждается, что качество
движения зависит от значения суммы (Da + t)
= Т. Этот закон был, в принципе, подтвержден рядом исследователей для Da ≈ < 100 мс
[6], [42], [50], [51], [60], но существуют разногласия по поводу вида функции,
связывающей Da с t.
Считается, что это, во всяком случае, не линейная функция, как это представлялось
Корте.
Правда, Нейгауз сделал справедливое замечание, что бета-движение при
условии Da = 10 мс, t = 90 мс заметно отличается от бета-движения при условии Da = 90 мс, t== 10 мс [51]. Однако в обоих случаях,
независимо от величины Da, оптимальным значением Т остается 100 мс.
Феноменальные же отличия названы меньшей видимой яркостью фигуры в первом
случае и тем, что во втором случае значение Da близко к порогу «неподвижности».
Большую стабильность показывает Т также при изменении α [6], [45], [51], что противоречит третьему закону
Корте, устанавливающему прямо пропорциональные отношения между t и α.
Однако очевидно, что при больших Da (например, 5 с или больше) качество бета-движения зависит только от t. Канеман и Уолман считают, что если Da ≈ < 100 мс, то
бета-движение зависит от Т, а если Da ≈ > 100 мс, то от t
[42], [43]. Обе эти зависимости объединяются авторами в законе интервала между
окончанием реакции на фигуру а и началом реакции на фигуру b (in-terresponse interval—IRI): качество видимого движения зависит только
от значения IRI для
данного стробоскопического стимула. IRI,
соответствующий оптимальному движению, является отрицательной величиной и
константен при всех значениях времени экспозиции обеих фигур [43, 161]. В
основе закона IRI
лежит предположение о том, что феноменальная продолжительность вспышек при Da ≈ < 100 мс
постоянна, а при Da ≈
> 100 мс прямо пропорциональна величине Da. Поэтому при изменении Da в
диапазоне 0<Da ≈
< l00 мс значение IRI будет оставаться постоянным, пока Т = const. Когда Da изменяется в диапазоне 100 мс ≈ < Da <
∞, значение IRI
будет оставаться постоянным, если теперь уже t = const. По мнению авторов, наилучшие условия для бета-движения
будут тогда, когда начало разрушения образа фигуры а совпадает с началом построения
образа фигуры b.
Закон IRI можно считать весомым
вкладом в решение проблемы взаимосвязи восприятия
135
формы и
видимого движения, которая является одной из основных в рамках исследования
видимого движения [6], [131, [22], [25], [34], [45], [46], [53], [65].
Одна из гипотез на этот счет была предложена Б. М. Величковским
и его сотрудниками [1], [2], [3]. Если появление в зрительной системе реакции
на фигуру b застает образ фигуры а
на раннем этапе его построения (при коротких Т), то образ фигуры а, вовлекаясь
в движение, поддается пластичному изменению (при диспаратности
форм a и b). При этом незавершенность построения образа
фигуры а может привести к некоторому ухудшению восприятия формы или текстуры
фигуры [2], [9], [23], [24], [25]. При больших Т микрогенез
формы уже завершен. В этом случае восприятие движения происходит на другом —
более высоком уровне, который обеспечивает восприятие инвариантной формы при
движении. Эта гипотеза, однако, испытывает трудность перед тем фактом, что
переход от пластичного изменения формы к движению ригидной формы при Da ≈ > 100 мс
зависит не от Т, а именно от t
[45], [46]. Если, например, Da = 400 мс, a t = 10 мс, то в 80 % случаев наблюдается
пластичное изменение формы [46], хотя микрогенез
формы уже полностью завершен [1]. С учетом данного факта более правомерным
будет предположение, что переход от одного типа движения к другому зависит не
от этапа построения образа фигуры а, но, наоборот, от этапа его разрушения.
Межстимульное
расстояние может изменяться
в широких пределах: от 2' [32] до, приблизительно, 4° [45]. Чем меньше α, тем больше диапазон изменения t, в котором наблюдается бета-движение [66].
В последнее время появились основания полагать, что процесс восприятия
движения при α < 15' отличается от процесса
восприятия движения при α ≈ < 15'.
Основания для выделения короткоамплитудного видимого
движения были подсказаны следующими остроумными экспериментами. В качестве
паттернов используются случайноточечные стереограммы, состоящие из черных и белых точек (тех и других
по 50 %), предъявляемые на одном месте. Распределение точек на обоих паттернах
не коррелировано, за исключением центральных зон,
имеющих обычно форму квадрата и смещенных относительно друг друга не более чем
на 15'. При рассматривании обоих паттернов вместе или поочередно наблюдатель не
в состоянии заметить идентичность центральных квадратов и вообще не может
выделить их из фона, т. е. попросту он их не видит. Однако, когда такие
паттерны предъявлялись монокулярно в стробоскопическом режиме, возникало
видимое движение центрального квадрата [20], [41], [49].
На первый взгляд видимое движение при таких условиях стимуляции можно
отождествить с уже описанным движением группы. Однако это будет неверно. Было
показано, что условия возникновения этого движения, напротив, больше
соответствуют условиям возникновения движения элемента. Основные факты
заключаются в том, что оба типа движения возникают при коротких t и не возникают при дихоптическом предъявлении
паттернов [54], [56].
Энстис считает, что движение коррелированных зон в случайноточечных
паттернах возникает в результате поточечного сравнения распределения яркостей
на паттернах а и b [12].
Если зрительный угол между соответственными точками в паттернах не превышает
15', то каждая пара таких точек представляет собой локальный стробоскопический
стимул. В пределах данного угла каждой точке центрального квадрата паттерна а
соответствуют несколько точек паттерна b, поэтому локальный стробоскопический стимул
представляет собой, очевидно, не тип а, b, а тип а, nb. Если бы оба паттерна были
некоррелированными по всей площади, то каждая точка паттерна а совершала бы
движение во всех возможных направлениях одновременно (эффект расщепления). В
целом такой хаос разнонаправленных движений трудно расценить как движение,
скорее, это похоже на мерцание. Именно это и наблюдается в некоррелированной
зоне случайноточечных паттернов. В коррелированной
зоне все точки, помимо случайных пар, имеют общую «судьбу», т. е. направление
движения и амплитуду. Таким образом, «индивидуальные» движения каждой точки
объединяются в движение всего центрального квадрата, который разделяет общую
«судьбу» всех точек. Восприятие формы коррелированных зон здесь вторично по
отношению к восприятию движения отдельных точек. Поэтому, как считает Брэддик, восприятие самой по себе формы коррелированных зон
может служить индикатором восприятия короткоамплитудного
движения [20]. Можно полагать, что α ≈ 15'
является пороговым для восприятия движения коррелированных зон в случайноточечных паттернах. Если а превышает это значение,
более выраженными оказываются случайные движения в других направлениях, а
движение центрального квадрата маскируется.
Петерсик и Пантл [56] считают, что восприятие короткоамплитудного движения (а также движения элемента а1—b3 (рис. 3, и) и движения изменяющейся формы) реализуется процессом,
который основан на отыскании локального соответствия элементов паттернов а и b — эпсилон-процессом.
Восприятие движения группы и движения ригидной формы опосредовано ипсилон-процессом, которому предшествует установление
глобального соответствия паттернов. Что представляют собой оба процесса, авторы
не уточняют.
Брэддик предлагает другую классификацию: восприятие короткоамплитудного
видимого движения представляет собой нейрофизиологический процесс, а восприятие
классического видимого движения (α ≈ >
15')—когнитивный [20], [21]. Ссылаясь на некоторые нейрофизиологические данные
[16], [35], Брэддик считает, что восприятие короткоамплитудного видимого движения так же, как и
реального, может обеспечиваться чувствительными к движению элементами сетчатки,
например ганглиозными клетками. Стробоскопический
стимул при α ≈ < 15' входит в диапазон
толерантности этих детекторов движения [4]. В пользу этой гипотезы говорят
следующие факты. Наблюдатели не могут отличить реальное
136
от видимого
движения, если α < 35'10" (Da = 50
мс, t = 50 мс, V = = 5,8 град./с) [31]. Короткоамплитудное
видимое движение вызывает послеэффект [14], что может
быть вызвано адаптацией чувствительных к движению нейронов [15].
Наша точка зрения ближе к концепции Петерсика
и Пантла. Прежде всего, проблема восприятия движения
не может сводиться даже в некоторой ограниченной ситуации к работе детекторов
движения. Мы можем воспринимать или ложно воспринимать, или не воспринимать
движение перемещающегося или неподвижного объекта — все зависит от системы
отсчета, в которую его помещает наблюдатель [10]. Поэтому восприятие движения в
его психологическом аспекте следует рассматривать как систему перцептивных
операций, отвечающую задаче, включенной в деятельность наблюдателя. Но даже
если оставить в стороне проблему выбора и построения системы отсчета
наблюдателем, главная проблема видимого движения заключается в том, почему мы
видим один объект, перемещающийся из положения а в положение b, а не последовательность дискретных фаз
движения. Потому что реагируют детекторы движения? Во-первых, это не ответ по
существу. Во-вторых, обнаружение в зрительной системе нейронов, селективно
отвечающих на движение в определенном направлении и с определенной скоростью,
еще не доказывает того, что эти нейроны имеют отношение к восприятию движения.
Возможно, они действительно регистрируют движение относительно сетчатки или
относительно наблюдателя, но их функция ограничивается вызыванием
ориентировочной реакций.
Мы поддерживаем гипотезу о том, что при предъявлении стробоскопического
стимула в зрительной системе возникает перемещение фокуса возбуждения от фигуры
а к фигуре b как результат
низкочастотной пространственной фильтрации [27], протекающей с большой
скоростью [22], [25]. В зависимости от значений параметров стимула этот процесс
может достигать разной степени выраженности. Она максимальна при α ≤ 15', и в этом случае стробоскопический
стимул вызывает такой же эффект, как и реальное движение. (Это единственный
случай, когда можно говорить о почти полной идентичности механизмов восприятия
реального и видимого движения. Здесь различия сохраняются только вплоть до
уровня чувственной ткани [7] в нейрофизиологических механизмах образования
движущегося пятна). Она минимальна (имеет околопороговое
значение) при условиях, ведущих к восприятию фи-движения,
движения ригидной формы или эффекта Тернуса. Кстати,
условия для восприятия этих типов движения приблизительно совпадают. Поэтому
можно сказать, что мы не воспринимаем всех фаз поворота фигуры в пространстве,
или эффекта Тернуса. Это скорее подразумеваемое, а не
воспринимаемое движение. То, что действительно существует при этом на уровне
чувственной ткани, — диспаратные во времени и пространстве пятна а и b, образуемые за счет более медленной
высокочастотной пространственной фильтрации, и перемещение от а к b пятна околопороговой
интенсивности.
Из этого «материала» наблюдатель, имея перцептивную
задачу, данную ему в форме представления о порождаемом образе, строит тот или
иной образ: фи-движение или его разновидности,
движение ригидной формы, эффект Тернуса, или сукцессивное мелькание паттернов. Но все эти эффекты
движения обязаны своим происхождением воображению и перцептивному
интеллекту наблюдателя, они не имеют достаточной чувственной наполненности. В чувственной ткани они находят лишь
некоторую опору, каркас. Так можно представить себе вращение в пространстве
плоскости, проходящей через трех мух, кружащихся под лампочкой. Но эта
плоскость, не имеющая своей полной чувственной основы, будет оставаться весьма призрачным
перцептом.
Таким образом, при некотором режиме предъявления стробоскопического
стимула процесс пространственно-временной фильтрации в зрительной системе
приводит к образованию двух структур в чувственной ткани. Первая структура
служит сенсорной основой видимого движения, ее выраженность связана с законами
Корте и в конечном счете — с законом IRI;
вторая — восприятия независимых мельканий фигур. При приблизительном равенстве
выраженности обеих структур наблюдатель может произвольно выбирать одну из них
или комбинацию из них в качестве сенсорной основы образа (при реальном движении
такой выбор отсутствует). Так, например, смотря на витрину магазина, мы можем
переключать свое внимание или на то, что выставлено в витрине, или на стекло
витрины, или на отражение улицы в нем. Можно также построить комбинацию из этих
изображений. В широких возможностях комбинирования или структурирования
сенсорного материала, подчиненного перцептивной
задаче наблюдателя, и заключается секрет богатства феноменологии видимого
движения. При свободном рассматривании стимуляции наблюдатель может случайно
«выхватить» из чувственной ткани одну из возможных структур и уже затем,
удерживая ее, решать, на что это похоже. Настоящая концепция, разумеется,
нуждается в дальнейшей теоретической и экспериментальной разработке [9].
1. Величковский Б. М.
Зрительная память и модели переработки информации человеком.— Вопросы психологии,
1977, № 6, с. 49—61.
2. Величковский Б. М., Капица
М. С, Цзен Н. В. Опознание цифр при
стробоскопическом движении. — В сб.: Новое в психологии. М.: МГУ, 1975, с.
61—68.
3. Величковский Б. М., Цзен Н. В. Микроструктурный анализ восприятия формы и
стробоскопического движения. — Эргономика. Труды ВНИИТЭ. М., 1973, вып. 5, с. 37—51.
4. Грегори Р. Л.
Глаз и мозг: Психология зрительного восприятия. — М.: Прогресс, 1970, с.
101—130.
5. Грэхем Ч. X. Зрительное
восприятие. — В кн.: Стивене С. С. (ред.). Экспериментальная психология. М.: Иностранная
литература, 1963, т. 2, с. 476—484.
6. Колерc П. Иллюзии
движения. — В кн.: Восприятие. Механизмы и модели. М., 1974, с. 309—322.
7. Леонтьев А. Н. Психология образа. — Вестник
Московского университета. Сер. 14.
137
Психология, 1979, № 2, с. 3—13.
8. Логвиненко А. Д., Миртов
Ю. Н. Видимое движение и метаконтраст: две стороны
одного явления. — Вестник Московского университета. Сер. 14. Психология, 1980, №
2, с. 37—47.
9. Миртов Ю. Н.
Маскировка при стробоскопической стимуляции. — Готовится к печати.
10. Рок И. Введение
в зрительное восприятие. — М.: Педагогика, 1980, кн. 1, с. 203—266.
11. Alpern M. Relation
of visual latency to intensity. — AMA Arch. Ophthal, 1954,
v. 51, p. 369—374.
12. Anstis S. M. Phi movement as a subtraction process.
— Vision Research, 1970, v. 10, p. 1411 — 1430 .
13. Anstis S. M.
Apparent movement. — In: R. Held, H. W. Leibowitz &
H. — L. Teuber (Eds.) Handbook of Sensory Physiology
(v. VIII) Perception.
14. Banks W. P., Kane D. A. Discontinuity
of seen motion reduces the visual motion aftereffect.—Perception and Psychophysics,
1972, v. 12, p. 69—72.
15. Barlow H. В.,
Hill R. M. Evidence for a physiological explanation of the waterfall phenomenon and
figural aftereffects. — Nature, 1963, v. 200, p. 1345—1347.
16. Barlow H. В., Levick W. R. The mechanism of directionally selective
units in the rabbit retina. — J. Physiol.,
17. Bartley S. H., Miller J. W. Some
circumstances surrounding apparent movement in the line of regard. — J. Psychol., 1954, v. 38, p. 453—456.
18. Bernstein A. S., Taylor K., Austen B. G., Nathanson N., Scarpelli A. Orienting
response and apparent movement toward or away from the observer.— J. Exp. Psychol., 1971, v. 87, p. 37—45.
19. Biederman-Thorson M., Thorson J., Lange G. D.
Apparent movement due to closely spaced sequentially flashed dots in the human
peripheral field of vision. — Vision Research., 1971, v. 11, p. 889—903.
20. Braddick O. A short-range process in apparent motion.
— Vision Research, 1974, v. 14, p. 519—527.
21. Braddick O., Adlard A.
Apparent motion and the motion detector. — In: Visual Psycho-physics and Physiology.
Armington J. C, Kxaskopf
J., Wooten B. R. (Eds.), N. Y.; Academic Press, 1978, p. 417—426.
22. Breitmeyer B. G. Unmasking visual masking: A look at the
"Why" behind the veil of the "How". — Psychol.
Review, 1980, v. 87, p. 52—69.
23. Breitmeyer В.,
Love R., Wepman B. Contour suppression during stroboscopic
motion and metacontrast. — Vision Research, 1974, v.
14, p. 1451—1456.
24. Breitmeyer В., Battaglia F., Weber С U-shaped backward contour
masking during stroboscopic motion. — J. Exp. Psychol.:
Human Perception and Performance, 1976, v. 2, p. 167— 173.
25. Breitmeyer B.
G., Ganz L. Implication of sustained and
transient channels for theories of visual pattern masking, saccadic suppression
and information processing. — Psychol. Rev., 1976, v.
83, p. 1—36.
26. Brown J. F., Voth
A. C. The path of seen movement as a function of the vector-field. — Amer.
J. Psychol., 1937, v. 49, p. 543—563.
27. Caelli T. M., Finlay
D. С Frequency, phase and colour coding in apparent motion. — Rerception,
1979, v. 8, p. 59—68.
28. Cohen J., Hansel C. E. M., Sylvester 1. D.
A new phenomenon in time judgement. — Nature,
1953, v. 172, p. 901.
29. DeSilva H. R. An experimental investigation of
the determinants of apparent visual movement. — Amer. J. Psychol.,
1926, v. 37, p. 469—501.
30. DeSilva H. R.
Kinematographic movement of parallel lines. — J. General
Psychol., 1928, v. 1, p. 550—577.
31. DeSilva H. R. An analysis of the visual
perception of movement. — Brit. J. Psychol., 1929, v.
19, p. 268—305.
32. Exner S. Ober das
Sehen von Bewegun-gen und
die Theorie das zusammengesetzen An-ges. — Sitzungsberichte Akademie Wissenschaft Wien, 1975, B. 72,
S. 156—190.
33. Geldreich E. W. A lecture room demonstration of the visual tau-effect. — Amer. J. Psychol.,
1934, v. 46, p. 483—485.
34. Griinau M. W., von. Dissotiation
and interaction of form and motion information in the human visual system. —
Vision Research, 1978, v. 18, p. 1485—1489.
35. Grusser O.J.,
36. Hall К R.
L., Earle A. E., Crookes T. G. A pendulum
phenomenon in the visual perception of apparent movement. — Quart. J. Exp. Psychol., 1952, v. 4. p. 109—120.
37. Higginson G. D. Apparent visual movement and Gestalt. I. Nine
observations which stand against Wertheimer's cortical
theory. — J. Exp. Psychol., 1926, v. 9, p. 228—252.
38. Home E. P., Parker R. Varying interstimulus distance and size of stimulus in beta motion.
— J. General Psychol., 1978, v. 99, p. 61—66.
39. Hovey H. B. Some factors influencing the brilliance
limen of vision. — Amer. J. Psychol.,
1936, v. 48, p. 434—445.
40. Johansson G. Configurations in Event
Perception.
41. lulesz B. Foundations
of Cyclopean Perception.
42. Kahneman D.
An onset-onset law for one case of apparent motion and metacontrast.
— Perception and Psychophysics, 1967, v. 2, p. 577—584.
43. Kahneman D., Wolman R. E. Stroboscopic motion:
effects of duration and interval. — Perception and Psychophysics, 1970, v. 8,
p. 161 — 164.
44. Kenkel F. Untersuchungen über den Zusammenhang swischen Erscheinungsgrasse und Erscheinungsbewegung
bei einigen sogenennten
138
optischen Tauschungen. — Zeits. P. Psychol., 1913, B. 67, S. 358—450.
45. Kolers P. A. Aspects of Motion Perception.
46. Kolers P. A.,
47. Korte A. Kinematoskopische Untersuchungen.
— Zeitz. f. Psychol., 1915,
B. 72, S. 193— 296.
48. Lane W. P., Home E. P. Stimulus
intensity and position effects on apparent movement. — J. General Psychol., 1964, v. 71, p. 87—92.
49. Lee D. N. Stimulus pairing in sequential
phi motion. — Perception, 1972, v. 1, p. 85—91.
50. Mizusava K,
51. Neuhaus W. Experimented Untersuchung
der Scheinbewegung. — Arch.
f. d. ges. Psychol., 1930,
B. 75, S. 315—458.
52. Newman. E. B. Versuche
über das Gamma-Phänomen. — Psychologische Forschung, 1934, B. 19, S. 102—121.
53. Orlansky I. The effect of similarity and difference in form
on apparent visual movement—Arch. Psychol., N.Y.,
1940, No. 246.
54. Pantle A., Picciano L. A
multistable movement display: Evidence for two
separate motion systems in human vision. — Science, 1976, v. 193, p. 500—502.
55. Petersik J. T. Two types of apparent movement in
the same visual display. — Paper presented at the 16-th annual meeting of the Psychonomic Society,
56. Petersik J. Т., Pantle A. Factors controlling the competing sensations
produced by a bistable stroboscopic motion display. —
Vision Research, 1979, v. 19, p. 143—154.
57. Pollack R. H. Initial stimulus duration
and the temporal range of apparent movement. — Psychonomic
Science, 1966, v. 5, p. 165—166.
58. Robins C, Shepard R. N. Spatio-temporal probing of apparent rotational movement. —
Perception and Psycho physics, 1977, v. 22, p. 12—18.
59. Saucer R. T. The nature of perceptual
processes. — Science, 1953, v. 117, p. 556—558.
60. Sgro F. J. Beta
motion thresholds. — J. Exp. Psychol., 1963, v. 66,
p. 281—285.
61. Stoper A., Bantly S.
Relation of split apparent motion to metacontrast. —
J. Exp. Psychol., Human Perception and Performance,
1977, v. 3, p. 258—277.
62. Ternus J. Experimented Untersuchungen
fiber phanomenale Identitat.
— Psychologische Forschung,
1926, B. 7, S. 81—136.
63. Tyler C. W. Temporal characteristics in
apparent movement: Omega movement vs. Phi movement. — Quart. J. Exp. Psychol., 1973, v. 25, p. 182—192.
64. Waals H. G., Roelofs С. О. Optische Scheinbewegung. — Zeits. f. Psychol., 1930, B. 114,
S. 241—288; B. 115, S. 91 — 193.
65. Warren W. H. Visual information for
object identity in apparent movement. — Perception and Psychophysics, 1977, v.
21, p. 264— 268.
66. Wertheimer M. Experimented Studien fiber das Sehen von Bevegung. — Zeits. f. Psychol., 1912, B. 61,
S. 161—265.
67. Wienke R. E., Steedman W. С Apparent motion in geometric depth. — Human Factors,
1965, v. 7, p. 215—218.
68. Zapparoli G. C. Perception of simultaneous stroboscopic
movements in different directions and the Gestalt theory hypothesis of
physiological shortcircuiting. — Acta
Psychologica,
69. Zeeman W. P. C, Roelofs С. О. Some aspects of apparent motion. — Acta Psychologica,
Поступила в редакцию 2.VII. 1982 г.