106
ИЗУЧЕНИЕ РОЛИ ИНТЕРФЕРЕНЦИОННЫХ МЕХАНИЗМОВ ПАМЯТИ В ПРОЦЕССЕ АДАПТАЦИИ
В. В. КОЛЫШКИН
Адаптация представляет собой процесс постоянного приспособления организма к окружающей среде, который заключается в изменении определенных форм поведения, направленных на удовлетворение основных и необходимых жизненных его потребностей.
Перемещение организма в новые, контрастные климатогеографические условия вызывает необходимость включения других, более адаптированных программ реагирования. При этом, как правило, часто требуется быстрое создание (выработка) новых программ на основе следов информации, поступающей из окружающей среды, что обязательно происходит при участии процессов памяти [4], [5], [6], [10], [11], [13].
Поскольку поступление информации осуществляется непрерывно, обязательно происходит взаимодействие (интерференция) в мозгу вновь поступающей и уже поступившей информации.
Для экстренного создания новых адаптивных программ требуется быстрая оценка и выделение необходимой, значимой для организма информации, что обязательно происходит при участии эмоциогенных структур мозга. Целесообразно полагать, что при селекции информации, приводящей к избирательному использованию (воспроизведению) эмоционально значимой информации, определенную и важную роль играют процессы интерференции (как проактивной, так и ретроактивной).
Поэтому представляется интересным и целесообразным выявление возможной роли интерференции в механизмах памяти в процессе адаптации к различным климатогеографическим условиям. В мировой литературе почти не имеется исследований, опирающихся на одновременную регистрацию физиологических показателей, характеризующих функциональное состояние организма.
Задачей исследования, таким образом, являлась регистрация интерференции в процессах памяти в двух вариантах: при взаимодействии индифферентной информации с индифферентной, и при взаимодействии эмоционально значимой информации с индифферентной, а также попытка обнаружения некоторых возможных физиологических коррелятов интерференции в процессе адаптации.
МЕТОДИКА ИССЛЕДОВАНИЯ
Для исследования интерференции при взаимодействии двух потоков индифферентной информации использовалась известная методика, описанная Ц. Флоресом [14] (рис. 1). Испытуемые получали инструкцию запоминать все слова посписочно. Затем через каждые 3 мин предъявлялись 12 пар списков, состоящих из 14 слов (темп чтения — одно слово в с, интервал между списками каждой пары — 3—4 с). В случайном порядке давалась команда воспроизвести первый или второй список, но во всех случаях испытуемые воспроизводили по 6 раз оба списка. Воспроизведение во всех случаях и далее длилось 1 мин. В контрольных экспериментах предъявлялись по 6 раз одинарные списки, которые надо было воспроизвести сразу же после их предъявления (контроль на проактивную интерференцию — ПАИ) или через 15—16 с после него (контроль на ретроактивную интерференцию — РАИ) [7], [14]. Значение интерференции подсчитывалось по известной формуле (см. рис. 1).
Для изучения интерференции эмоциональных стимулов на индифферентные была использована разработанная и опробованная нами методика. Испытуемым предъявлялись 12 раз одинарные списки из 14 индифферентных слов (темп чтения и интервал между списками тот же, что и в первом случае), но в середину шести списков включались 2 слова, подобранные путем независимой экспертной оценки и вызывающие яркие эмоциональные образы (порядковый номер слов — 7 и 8). Вследствие этого образная позиционная кривая [15] (кривая эффективности воспроизведения слов в зависимости от их места в описке) имела в своей середине значительный подъем (с 29—33 до 64—69%), в результате чего воспроизведение первых и последних шести слов в списке, как показали наши исследования, изменяется по сравнению с так называемыми «чистыми» списками, где форма кривой остается без изменения. Имеются данные, что ПАИ более сильно сказывается в отношении запоминания элементов, находящихся в конце списка, а РАИ — в его начале [7], [9], [14], поэтому изменение воспроизведения первых и последних шести индифферентных слов списка можно объяснить интерференционным торможением (соответственно ретроактивным и проактивным) со стороны двух эмоциональных слов, включенных в его середину.
Таким образом, как нам кажется, предложенная методика является адекватной для изучения действия эмоциональной информации на индифферентную и, в конечном счете, для регистрации изменений интерференции Эмоциональных стимулов на индифферентные.
Эксперименты проведены на 8 практически здоровых испытуемых 20—28 лет в различных климатогеографических условиях: Новосибирск (май — июнь), Памир, высота 3600 м над уровнем моря (июль 1978 г.), и Камчатка, 5-часовой трансмеридианальный перелет, муссонный климат (август 1979 г.). Позиционная кривая по каждому списку строилась по всем испытуемым; таким образом, 100%-ное воспроизведение каждого порядкового слова выявлялось в том случае, когда это слово воспроизводилось всеми восемью испытуемыми, и т. д. Затем строилась общая кривая по всем шести спискам.
Для каждого экспериментального дня составлялись примерно равные по силе программы, но ни одно слово не повторялось дважды ни в одной из них. Небольшое индивидуальное различие списков нивелировалось большим количеством списков (48) в каждой программе.
В ходе опыта регистрировали ЭЭГ, КГР и двигательную активность. За 20 мин записи фона по трансформации в ЭЭГ и отсутствию двигательной активности можно было судить о времени бодрствования испытуемых. Для выявления существенных отклонений в показателях применялась статистическая обработка
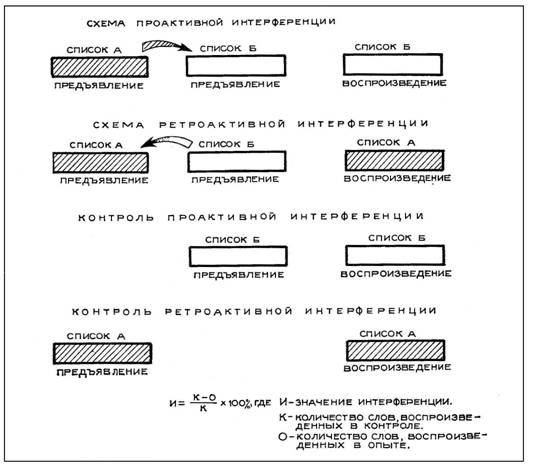
Рис. 1. Методика исследования интерференции.
усредненных результатов по группе на каждый день исследования. Каждая точка на графике представлена средней величиной по всем шести аналогичным значениям в программе и по всем восьми испытуемым. Достоверность отличий по отношению к исходным данным, полученным в Новосибирске, определялась с помощью парного критерия знаком [12] и критерия Вилкоксона [8].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ ОБСУЖДЕНИЕ
Трехдневные контрольные обследования в Новосибирске показали, что за 20 мин записи фона испытуемые находились в состоянии спокойного бодрствования 15—17 мин. За это время при регистрации ЭЭГ был отмечен выраженный альфа-ритм. Кратковременные эпизоды сна характеризовались отсутствием двигательной активности и изменением структуры ЭЭГ, свойственной В-стадии сна.
ПАИ и РАИ в Новосибирске перед памирской экспедицией достигали значений соответственно 24,2 и 63,3%, перед экспедицией на Камчатку—10,0 и 53,9% (среднее по двум дням обследования). Полученные количественные данные являются близкими величинами, при этом состав испытуемых в обеих экспедициях не повторялся.
Это соотношение ПАИ и РАИ сохраняется (РАИ больше ПАИ) и в случае взаимодействия эмоциональной и индифферентной информации. РАИ составляет 10,2%, а ПАИ имеет даже отрицательное значение (—0,9%), т. е. последние 6 слов в списке с включенными в середину двумя эмоциональными словами воспроизводятся даже несколько лучше, чем последние 6 слов в контрольных «чистых» списках.
Результаты двух экспедиционных исследований (Памир, 1978 г., и Камчатка, 1979 г.) показали однородные результаты. В случае гомогенной индифферентной информации ПАИ, увеличиваясь в первые дни адаптации, затем уменьшается на 4-й день и возвращается к норме к концу обследуемого периода (рис. 2(2)
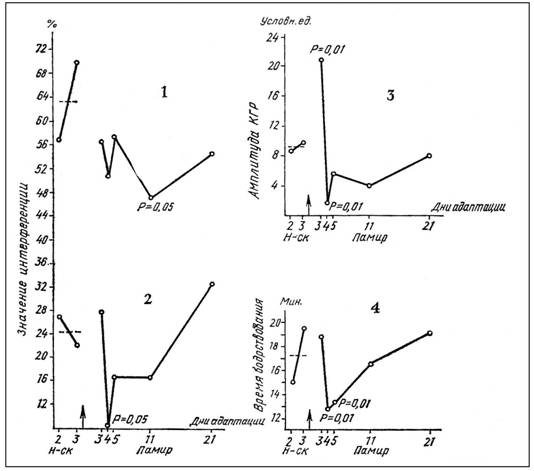
Рис. 2. Памир, ледник Шуровского, высота 3600 м над уровнем моря,
1— изменение ретроактивной интерференции
2— изменение проактивной интерференции,
3 — изменение фоновой кожногальванической реакции,
4 — изменение времени бодрствования.
109
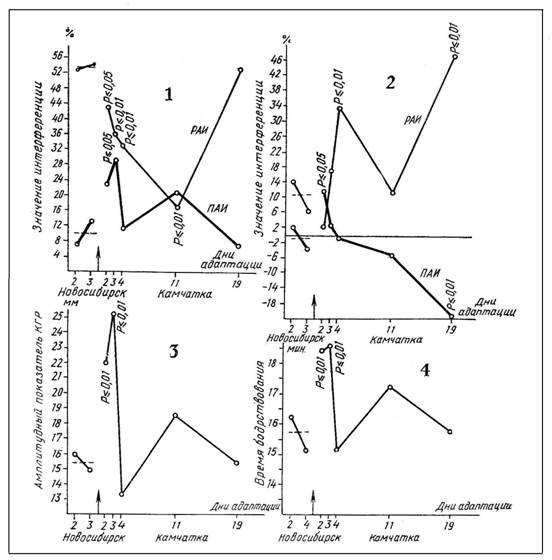
Рис. 3. Камчатка, п. Паратунка
1 — изменение интерференции индифферентных слов на индифферентные,
2 — изменение интерференции эмоциональных слов на индифферентные,
3 — изменение фоновой кожногальванической реакции,
4 — изменение времени бодрствования.
и 3(l)). РАИ остается в течение всего обследуемого времени ниже исходного уровня, зарегистрированного в Новосибирске (рис. 2(1) и 3(1)). Интересными являются общие вышеописанные закономерности изменений интерференции при исследовании обеих экспедиций, но формы соответствующих кривых, полученных при обследовании каждой из них, не совпадают, что объясняется, несомненно, действием совершенно различных по экстремальности климатогеографических условий.
В результате экспериментов выявлена сопряженность изменений ПАИ индифферентных слов на индифферентные, спонтанных КГР при записи фона и времени бодрствования (рис. 2 (2, 3, 4) и 3(1, 3, 4)), что говорит о роли активационных влияний ретикулярной формации (РФ) мозгового ствола в осуществлении процессов формирования следов памяти. Это согласуется с данными, полученными в лабораторных исследованиях С. П. Бочаровой и А. И. Лактионовым [2], которые обнаружили высокую корреляцию между уровнем ПАИ и лабильностью ЦНС (длительностью депрессии альфа-ритма и латентным периодом реакции десинхронизации). Изменения РАИ существенно отличны (рис. 2(1) и 3(1)). Аналогичные результаты, полученные в клинике [3], выявили значительно более выраженное положительное действие активационных влияний РФ на процессы формирования (консолидации) следа, чем на процессы его воспроизведения (репродукции).
Результаты исследований позволяют предложить следующую физиологическую трактовку этих различий. Интерференция — это взаимодействие
110
в мозгу следов памяти. В случае ПАИ след ранее поступившей информации влияет на формирование следа поступающей позднее информации. В случае же РАИ текущая информация воздействует на ранее сформированный след памяти, т. е., по-видимому, может влиять только на воспроизведение уже сформированного следа.
Изменения интерференции эмоциональных слов на индифферентнее значительно отличаются. Если РАИ индифферентных слов на индифферентные в течение всего процесса адаптации остается ниже исходного уровня, зарегистрированного в Новосибирске, то РАИ эмоциональных слов на индифферентные — выше этого уровня. Если ПАИ индифферентных слов на индифферентные возрастает, то при действии эмоциональных слов на индифферентные отмечено ее уменьшение. По-видимому, следует предположить различное влияние друг на друга эмоционально значимой и индифферентной информации.
Отмеченные 2 типа изменений интерференции обоих видов объясняются, по-видимому, следующим образом. Первый тип изменений (в случае гомогенной индифферентной информации) является общей схемой и характеризует принцип (механизм) взаимодействия информации вообще. Второй тип является частным случаем взаимодействия эмоциональной и индифферентной информации и выявляет возможный механизм выделения эмоционально значимой для организма информации.
Тогда становятся понятными возможный механизм выработки новых адаптивных программ и место интерференции в этом процессе. Уменьшение РАИ по первому типу, с одной стороны, создает условия для лучшего воспроизведения информации вообще, но, с другой стороны, уже в это же время в условиях острого цейтнота происходит отбор информации по критерию ее значимости [1]. Эмоционально значимая для организма информация подавляет поступление индифферентной за счет увеличения ПАИ (рис. 3(2). Это приводит к быстрому созданию основной конструкции новой программы взаимодействия организма с измененной внешней средой. В то же время воспроизведение индифферентной информации, поступившей ранее, подавляется в течение всего приспособительного периода вследствие роста РАИ со стороны эмоционально значимой информации.
Таким образом, уменьшение РАИ индифферентных стимулов на индифферентные создает условия для большего извлечения всей поступившей информации. Из всей этой информации уже в ранние сроки адаптации начинает выбираться эмоционально значимая. При этом РАИ подавляет воспроизведение индифферентной информации, а ПАИ — новое поступление. В более поздний период адаптации поступление индифферентной информации облегчается вследствие уменьшения ПАИ со стороны эмоциональных стимулов. Все это позволяет организму оперативно пользоваться не только старым, но и приобретенным, жизненно важным опытом и способствует ускоренной выработке адаптивного поведения.
На основании вышеизложенного можно сделать следующие выводы:
1. В случае однородной индифферентной информации обнаружены взаимосвязь изменений спонтанных КГР при записи фона и времени бодрствования с ходом проактивной интерференции и отсутствие видимой корреляции этих показателей с уровнем ретроактивной интерференции в процессе адаптации, что позволяет говорить о более значительной роли механизмов бодрствования в процессе формирования следа памяти по сравнению с его воспроизведением.
2. Получен факт разнонаправленного изменения интерференции эмоциональных стимулов на индифферентные и индифферентных друг на друга, что позволяет сделать предположение о различных механизмах этих интерференции.
3. Зарегистрированные изменения интерференции позволяют судить об общей универсальной роли интерференции в механизмах памяти в процессе адаптации к различным климатогеографическим условиям.
1. Бочарова С. П. Переработка знаковой информации в процессе памяти. — В кн.: Психологические проблемы переработки знаковой информации. М, 1976. — 140 с.
2. Бочарова С. П., Лактионов А. И. Интерференция в краткосрочной памяти и типологические свойства нервной системы человека. — В кн.: Материалы 4-го Всесоюзного съезда Общества психологов. Тбилиси, 1971, с. 385—386.
3. Вейн А. М., Каменещая Б. И. Память и функциональное состояние мозга. — Физиология человека, 1976, т. 2, № 3, с. 400—406.
4. Ильюченок Р. Ю. Фармакология поведения и памяти. — Новосибирск, 1971. — 223 с.
5. Ильюченок Р. Ю. Память и адаптация. — Новосибирск, 1979.— 192 с.
6. Кипиани Т. И., Сысоева А. Ф., Кулиева Э. А., Мегрелишвили С. И. Приспособительная деятельность центральной нервной системы при адаптации к климату Среднегорья. — В кн.: XXIV Всесоюзное совещание по проблемам высшей нервной деятельности. М., 1974. — 154 с.
7. Клацки Р. Память человека. Структура и процессы. М., 1978. — 319 с.
8. Лакин Г. Ф. Биометрия. — М., 1973. — 343 с.
9. Линдсей П., Норман Д. Переработка информации у человека. — М., 1974.
10. Меерсон Ф. З. Общие механизмы адаптации и профилактики. — М., 1973. — 360 с.
11. Миррахимов М. М., Соложенкин В. В., Шелухина Э. П., Мельников И. П., Новикова Н. Ф. Изучение психической работоспособности в процессе адаптации и реадаптации человека к условиям высокогорья Тянь-Шаня. — В кн.: Человек и среда. Л., 1975, с. 59—70.
12. Рокицкий П. Ф. Биологическая статистика. — Минск, 1973.— 97 с.
13. Слоним А. Д. Среда и поведение. Формирование адаптивного поведения. — Л., 1976. — 211 с.
14. Флорес П. Память. — В кн.: Экспериментальная психология / Под ред. П. Фресса и Ж- Пиаже. М., 1973, вып. 4, с. 209—342.
15. Murdock B.B., Jr. The serial position effect of free recall. - J. of Experimental Psychology, 1962, V. 64, р. 482—488.
Поступила в редакцию 16.ХI.1981 г.