44
ТЕОРЕТИЧЕСКИЕ ИССЛЕДОВАНИЯ
ПОТРЕБНОСТНО-ИНФОРМАЦИОННАЯ ТЕОРИЯ ЭМОЦИЙ
П. В. СИМОНОВ
I. ЧТО ТАКОЕ ЭМОЦИЯ?
Физиология конца XIX — начала XX столетия рассматривала эмоциональные реакции животных и человека преимущественно в связи с их отношением к вегетативно-гуморальным функциям организма. Эта тенденция отчетливо выступила в попытках определить роль висцеральных сдвигов в формировании эмоциональной окраски стимулов, действующих на организм (теория Джеймса — Ланге), равно как и в работах, рассматривавших эмоции в качестве состояний экстренной мобилизации вегетативных ресурсов (теория Кэннона — Барда). Однако начиная с конца 30-х гг. ученые все больше концентрируют свое внимание на тех факторах взаимодействия организма с внешней средой, которые ведут к возбуждению мозговых структур, ответственных за эмоциональные реакции. Эмоции и восприятие, эмоции и действия, эмоции и информация — вот те проблемы, которые заняли господствующее положение в современной психофизиологии эмоций. «Нужно думать,— говорил И. П. Павлов [8; 423] в 1932 г. с трибуны Международного конгресса в Риме,— что нервные процессы полушарий при установке и поддержке динамического стереотипа есть то, что обыкновенно называется чувствами в их двух основных категориях — положительной и отрицательной и в их огромной градации интенсивностей. Процессы установки стереотипа, довершения установки, поддержки стереотипа и нарушений его и есть субъективно разнообразные положительные и отрицательные чувства, что всегда и было видно в двигательных реакциях животного».
Эту павловскую идею несовпадения (рассогласования — скажем мы сегодня) заготовленного мозгом внутреннего стереотипа с изменившимся внешним мы не раз встретим в той или иной модификации у ряда авторов, обращавшихся к изучению эмоций. По мнению Хоуджа [23], эмоции возникают в момент, когда высшие мозговые центры не могут обеспечить адекватный ответ на воспринимаемую ситуацию или когда существуют сомнения, колебания относительно возможности успешного ответа. По Хоуджу, сила эмоциональной реакции обратно пропорциональна возможности высших центров мозга адекватно ответить на данную ситуацию. Эмоции представляют неудачу интеграции на уровне церебральной коры.
Близкие теоретические представления были позднее развиты Хеббом [22] на примере активации врожденного механизма страха. Согласно Хеббу, этот механизм вовлекается в процесс поведения, когда ситуация оказывается «странной» — частично знакомой, частично нет и не вполне понятной. Реакция страха основывается скорее на переживании рассогласования, чем на собственно сенсорном восприятии сложившейся обстановки.
«Биологическая теория эмоций» П. К. Анохина [1] непосредственно связана с его более общей теорией функциональной системы поведенческого акта. Согласно Анохину, нервный аппарат отрицательных и положительных эмоций активируется в тот момент, когда обнаруживается рассогласование или совпадение акцептора действий (афферентной модели ожидаемых результатов) с импульсацией, сигнализирующей о реально достигнутом эффекте. Мы завершим обзор этой линии исследований, органически близких идее И. П. Павлова, сформулированной им в 1932 г., ссылкой
45
на статью К. Прибрама [24] «Новая биология и неврология эмоций. Структурный подход», появившуюся в 1967 г. уже после того, как была опубликована [13] и доложена на XVIII Международном психологическом конгрессе в Москве «информационная теория эмоций» [14].
В 1924 г. советский психиатр В. П. Осипов [7] проницательно назвал первую стадию образования любого условного рефлекса — стадию генерализации — «эмоциональной» в отличие от более поздней «интеллектуальной, познавательной» стадии хорошо упроченного рефлекса. Эти две стадии, динамику ослабления, а затем и постепенного исчезновения эмоционального напряжения легко продемонстрировать на примере выработки оборонительного условного рефлекса у человека. Испытуемых просили нажимать на ключ через 20 с после короткого звукового сигнала. Если субъект нажимал на ключ раньше 19 с после подачи сигнала или позднее 21 с, ему наносили на кожу предплечья болевое раздражение электрическим током порядка 60—90 В. После каждой пробы субъекта информировали о времени его реакции. Степень эмоционального напряжения измеряли по изменению частоты сердцебиений. Нарастание частоты сердцебиений оценивали по суммарной продолжительности первых трех ударов сердца после звукового сигнала и последних трех ударов перед двигательной реакцией.
Если сравнить две стадии эксперимента, содержащие одинаковое число проб и одинаковое количество болевых раздражений током, можно убедиться, что суммарное отклонение частоты сердцебиений от исходного фона в этих двух частях эксперимента различно. Следовательно, оно зависит не только от количества наказаний. Опыты показали, что суммарное изменение частоты сердцебиений пропорционально суммарному отклонению времени двигательных реакций от заданной величины, т. е. пропорционально степени совершенства, точности и надежности условного оборонительного рефлекса. Это правило справедливо и для тех случаев, где величина ошибок (но не их количество, не количество болевых раздражений) нарастала вместе с нарастанием частоты сердцебиений и где, следовательно, динамику вегетативных сдвигов нельзя было объяснить привыканием к болевым стимулам по мере их повторения. Суммируя результаты собственных опытов и данные литературы, мы пришли в 1964 г. к выводу о том, что эмоция есть отражение мозгом человека и животных какой-либо актуальной потребности (ее качества и величины) и вероятности (возможности) ее удовлетворения, которую мозг оценивает на основе генетического и ранее приобретенного индивидуального опыта.
В самом общем виде правило возникновения эмоций можно представить в виде структурной формулы:
Э = f [П, (Ин –– Ис,...], где
Э — эмоция, ее степень, качество и знак; П — сила и качество актуальной потребности; (Ин — Ис) — оценка вероятности (возможности) удовлетворения потребности на основе врожденного и онтогенетического опыта; Ин — информация о средствах, ресурсах и времени, прогностически необходимых для удовлетворения потребности; Ис — информация о средствах, ресурсах и времени, которыми располагает субъект в данный момент.
Разумеется, эмоция зависит и от ряда других факторов, одни из которых нам хорошо известны, а о существовании других мы, возможно, еще и не подозреваем. К числу известных относятся:
индивидуальные (типологические) особенности субъекта, прежде всего индивидуальные особенности его эмоциональности, мотивационной сферы, волевых качеств и т.п.;
фактор времени, в зависимости от которого эмоциональная реакция приобретает характер стремительно развивающегося аффекта или настроения, сохраняющегося часами, днями и неделями;
качественные особенности потребности. Так, эмоции, возникающие на базе социальных и духовных потребностей, принято именовать чувствами. Низкая вероятность избегания нежелательного воздействия породит у субъекта тревогу, а низкая вероятность достижения желаемой цели — фрустрацию и т.д. и т.п.
Информационная теория эмоций справедлива не только для сравнительно сложных поведенческих и психических актов, но для генеза любого эмоционального состояния. Например, положительная эмоция при еде возникает за счет интеграции голодового возбуждения (потребность) с афферентацией из полости рта, свидетельствующей о растущей вероятности удовлетворения данной потребности. При ином состоянии потребности та же афферентация окажется эмоционально безразличной или генерирует чувство отвращения.
До сих пор мы говорили об отражательной функции эмоций, которая совпадает с их оценочной функцией. Обратите внимание, что цена, в самом общем смысле этого понятия, всегда есть функция двух факторов: спроса (потребности) и предложения (возможности эту потребность удовлетворить). Но категория ценности и функция оценивания становятся ненужными, если отсутствует необходимость сравнения, обмена, т. е. необходимость сопоставления ценностей. Вот почему функция эмоций не сводится к простому сигнализированию воздействий полезных или вредных для организма, как полагают сторонники «биологической теории эмоций». Воспользуемся примером, который приводит П. К. Анохин [1; 342]. При повреждении сустава чувство боли ограничивает двигательную активность конечности, способствуя репаративным процессам. В этом интегральном сигнализировании «вредности» П. К. Анохин видел приспособительное значение боли. Однако аналогичную роль мог бы играть механизм, автоматически, без участия эмоций, тормозящий движения, вредные для поврежденного органа. Чувство боли оказывается более пластичным механизмом: когда потребность в движении становится очень велика (например, при угрозе самому существованию субъекта), движение осуществляется, невзирая на боль. Иными словами, эмоции выступают в роли своеобразной «валюты мозга» — универсальной меры ценностей, а непростого эквивалента, функционирующего по принципу: вредно — неприятно, полезно — приятно.
Из существа отражательно-оценочной функции эмоций вытекают их регуляторные функции. Мы начнем их анализ с функцией, регулирующих индивидуальное поведение.
Переключающая функция эмоций. С физиологической точки зрения, эмоция есть активное состояние системы специализированных мозговых структур, побуждающее изменить поведение в направлении минимизации или максимизации этого состояния. Поскольку положительная эмоция свидетельствует о приближении удовлетворения потребности, а отрицательная эмоция — об удалении от него, субъект стремится максимизировать (усилить, продлить, повторить) первое состояние и минимизировать (ослабить, прервать, предотвратить) второе. Этот гедонистический принцип максимизации — минимизации, равно применимый к человеку и животным, позволяет преодолеть кажущуюся недоступность для непосредственного экспериментального изучения эмоций животных.
Переключающая функция эмоций обнаруживается как в сфере врожденных форм поведения, так и при осуществлении условнорефлекторной деятельности, включая ее наиболее сложные проявления. Надо лишь помнить, что оценка вероятности удовлетворения потребности может происходить у человека не только на осознаваемом, но и на неосознаваемом уровне. Ярким примером неосознаваемого прогнозирования служит интуиция, где оценка приближения к цели или удаления от нее первоначально реализуется в виде эмоционального «предчувствия решения», побуждающего к логическому анализу ситуации, породившей эту эмоцию.
Переключающая функция эмоций особенно ярко обнаруживается в процессе конкуренции мотивов, при выделении доминирующей потребности, которая становится вектором целенаправленного поведения. Так, в боевой обстановке борьба между естественным для человека инстинктом самосохранения и социальной потребностью следовать определенной этической норме
47
переживается субъектом в форме борьбы между страхом и чувством долга, между страхом и стыдом. Зависимость эмоций не только от величины потребности, но и от вероятности ее удовлетворения чрезвычайно усложняет конкуренцию сосуществующих мотивов, в результате чего поведение нередко оказывается переориентированным на менее важную, но легко достижимую цель: «синица в руках» побеждает «журавля в небе». Постоянная угроза такой переориентации потребовала от эволюции формирования специального мозгового аппарата, способного купировать эту «ахиллесову пяту» эмоций. На уровне высшей нервной деятельности человека он предстает перед нами в качестве физиологических механизмов воли.
Частной, хотя и качественно специфичной разновидностью переключающей функции эмоций является их подкрепляющая функция.
Роль эмоций в замыкании условного рефлекса мы пытались схематически изобразить на рис. 1. С момента своего возникновения условнорефлекторная теория предполагала конвергенцию двух возбуждений: от условного раздражителя и от стимула, вызывающего безусловный рефлекс, например афферентацию из полости рта при поступлении в рот пищи (I на рис. 1). Тогда же выяснилось значение «наличного функционального состояния», пищевой возбудимости, которую сегодня мы можем рассматривать как результат возбуждения мозговых структур, активированных возникновением соответствующей потребности, состоянием голода (II на рис. 1).
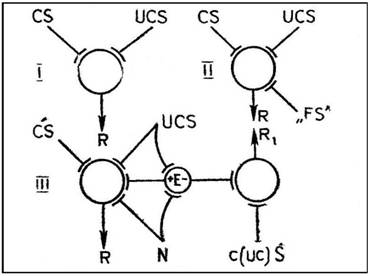
Рис. 1. Двух- (I), трех- (II) и четырехфакторные (III) схемы встречи возбуждений в пункте конвергенции при формировании условного рефлекса.
–– условный стимул, —безусловный стимул, — реакция,
–– функциональное состояние, — потребность, — эмоция
Однако ни афферентация из полости рта, ни голодовое возбуждение сами по себе не могут играть роль подкрепления, обеспечивающего формирование инструментального условного рефлекса. Только интеграция голодового возбуждения с возбуждением от фактора, способного удовлетворить данную потребность, т. е. механизм, генерирующий положительную эмоцию, обеспечивает выработку условного рефлекса. При ином соотношении конвергирующих возбуждений, например при поступлении пищи в рот перекормленного животного, активация механизмов отрицательной эмоции приведет к оборонительной реакции избегания (III на рис. 1).
Участие нервных механизмов эмоций в процессе выработки любого инструментального рефлекса делает весьма относительным выделение феномена так называемой эмоциональной памяти. По-видимому, мы можем говорить только о большей или меньшей силе эмоциональной реакции на подкрепляющий стимул, как это имеет место при выработке условного рефлекса после одного сочетания или при выработке условной пищевой аверсии, когда подкрепление (отравление животного) происходит через несколько часов после восприятия условного сигнала. Что касается эмоций в момент воспроизведения условных связей, то они зависят от степени актуальности той потребности, на базе которой возникает данная эмоциональная реакция. Экспериментально показано [16; 34], что дети 8 лет через 10 дней после первого опыта лучше воспроизводили тот словесный материал, который соответствовал мотиву, доминирующему в их личностной иерархии, будь то враждебность, лидерство, любознательность, привязанность и т. д.
Экспериментально установленная роль эмоций в процессе выработки классических и особенно инструментальных условных рефлексов позволяет утверждать, что без учета и дальнейшей разработки нейрофизиологии
48
эмоций мы вряд ли решим вопрос о механизмах замыкания условных связей.
Компенсаторная (замещающая) функция эмоций. Будучи активным состоянием системы специализированных мозговых структур, эмоции оказывают влияние на другие церебральные системы, регулирующие поведение, процессы восприятия внешних сигналов и извлечения энграмм этих сигналов из памяти, вегетативные функции организма.
Но компенсаторная функция эмоций отнюдь не ограничивается гипермобилизацией вегетатики. Возникновение эмоционального напряжения сопровождается переходом к иным, чем в спокойном состоянии, формам поведения: иным принципам оценки внешних сигналов и иному реагированию на них. Физиологически суть этого перехода можно определить как возврат от тонко специализированных условных реакций к реагированию по принципу доминанты А. А. Ухтомского.
Эти переходы подчиняются универсальному для головного мозга «закону обратных отношений между рефлексами», сформулированному А. А. Ухтомским [18; 246]. В наиболее сложных и высших своих проявлениях этот универсальный закон обнаруживается в том, что «между доминантой (внутренним состоянием) и данным рецептивным содержанием (комплексом раздражителей) устанавливается прочная («адекватная») связь, так что каждый из контрагентов (внутреннее состояние и внешний образ) будут вызывать и подкреплять исключительно друг друга» [17; 169]. Эта закономерность подверглась анализу в трудах И. С. Беритова и И. П. Павлова, она была всесторонне исследована Э. А. Асратяном [2], [3] на примере взаимодействия прямых и обратных условных связей.
Совершенно очевидно, что предположительное доминантное реагирование целесообразно только в условиях прагматической неопределенности. При устранении этой неопределенности субъект может превратиться в «пуганую ворону, которая и куста боится». Вот почему эволюция сформировала механизм зависимости эмоционального напряжения и характерного для него типа реагирования от размеров дефицита прагматической информации, механизм элиминирования отрицательных эмоций по мере ликвидации информационного дефицита. Подчеркнем, что эмоция сама по себе не несет достаточной информации об окружающем мире, недостающая информация пополняется путем поискового поведения, совершенствования навыков, мобилизации хранящихся в памяти энграмм. Компенсаторное значение эмоций заключается в их замещающей роли.
Что касается положительных эмоций, то их компенсаторная функция реализуется через влияние на потребность, инициирующую поведение. В трудной ситуации с низкой вероятностью достижения цели даже небольшой успех (возрастание вероятности) порождает положительную эмоцию воодушевления, которая усиливает потребность достижения цели согласно правилу
П = Э/(Ин –– Ис),
вытекающему из формулы эмоций.
В иных ситуациях положительные эмоции побуждают живые существа нарушать достигнутое «уравновешивание с окружающей средой». Стремясь к повторному переживанию положительных эмоций, живые системы вынуждены активно искать неудовлетворенные потребности и ситуацию неопределенности, где полученная информация могла бы превысить ранее имевшийся прогноз. Тем самым положительные эмоции компенсируют недостаток неудовлетворенных потребностей и прагматической неопределенности, способных привести к застою, к деградации, стать препятствием для процесса самодвижения и саморазвития. Наличие отрицательных и положительных эмоций свидетельствует о дифференциации потребностей на две основные категории: потребности нужды и потребности роста, которым в физиологии соответствуют представления о побуждении и желании, о двух разновидностях мотиваций — отрицательных и положительных (аппетит).
До сих пор мы говорили об участии эмоций в организации индивидуального
49
поведения. Не меньшее значение эмоции имеют для поведения группы, сообщества, колонии. Здесь также обнаруживаются все три регуляторные функции эмоций: переключающая, подкрепляющая («эмоциональный резонанс») и компенсаторная (подражательное поведение). Подробнее об этом — в нашей книге «Эмоциональный мозг» [16].
Классификация эмоций. Многообразие потребностей, сосуществующих и образующих сложные, иерархически организованные системы, делает составление сколько-нибудь «полных», «развернутых» классификаций эмоций делом абсолютно бессмысленным и безнадежным. Вот почему большинство авторов пытается определить ограниченное число базальных эмоций, не удовлетворяясь их делением только на положительные и отрицательные.
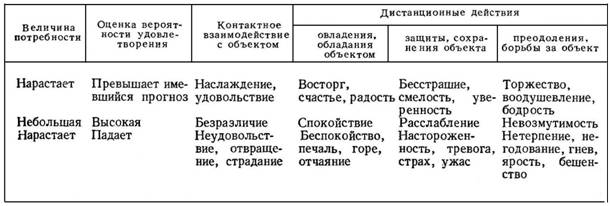
Поскольку вероятность удовлетворения потребностей в огромной мере зависит от действий субъекта, мы предположили, что именно характер действий может служить классифицирующим принципом для выявления фундаментальных эмоций, занимающих центральное положение в сфере эмоциональных состояний человека [15]. Взаимодействие с объектом удовлетворения потребности, в свою очередь, бывает либо контактным, которое субъект может прервать или продолжить, но не предотвратить, либо дистанционным. Что касается дистанционных действий, то, согласно военной науке, они существуют в трехосновных разновидностях: наступление (преодоление), оборона (защита, сохранение) и отступление (утрата ранее занимаемых позиций). Эмоции, соответствующие перечисленным типам взаимодействия, представлены в табл. 1. В случае одновременной актуализации двух и более потребностей они могут порождать сложнейшие эмоциональные аккорды.
Таблица 1
КЛАССИФИКАЦИЯ ЭМОЦИЙ В ЗАВИСИМОСТИ ОТ ВЕЛИЧИНЫ ПОТРЕБНОСТИ, ВЕРОЯТНОСТИ ЕЕ УДОВЛЕТВОРЕНИЯ И ХАРАКТЕРА ДЕЙСТВИЙ
Помимо характера действий происхождение базисных эмоций можно связать и с тремя основными группами потребностей: биологических, социальных и идеальных. Тогда биологическим потребностям самосохранения (в широком смысле) будет соответствовать страх. Гнев возникает в сфере внутривидового взаимодействия, вторично распространяясь на неодушевленные предметы. Человек может прийти в ярость, преодолевая преграду из камней или пытаясь починить испорченный мотор, хотя злиться на камни или машину нелепо. Что касается положительных эмоций типа радости, то они впервые возникают у ребенка в процессе познания окружающего мира (формирование механизмов ясного видения при конвергенции зрительных осей) и первичного общения с другими людьми. Удовлетворение биологических потребностей (голод, температурный дискомфорт) внешних признаков положительных эмоций у ребенка не вызывает: младенец просто успокаивается и засыпает. Эмоции удовольствия и отвращения возникают при контактном взаимодействии в связи с удовлетворением любой потребности: мы можем наслаждаться и
50
вкусной пищей, и созерцанием произведения искусства, хотя, разумеется, это очень разные типы наслаждения. Пока что трудно ответить на вопрос, какому фактору принадлежит решающая роль в происхождении трех фундаментальных эмоций: характеру действий или трем основным классам потребностей. Вполне возможно, что в процессе эволюции сыграли свою роль оба фактора: действия и побуждавшие их потребности.
II. ЭМОЦИОНАЛЬНЫЙ МОЗГ
Понимая обоснованность разочарования в результатах поиска «точечно» локализованных эмоциогенных структур, мы вместе с тем не можем вернуться к расплывчатым представлениям об эмоциях как функции «мозга в целом», как следствия «корково-подкорковых отношений». При всей сложности проблемы экспериментальная нейрофизиология эмоций не вправе отказаться от фундаментального принципа «приурочивания динамики к структуре», завещанного нам И. П. Павловым.
Сегодня уже невозможно найти исследователя, который представлял бы себе нервные центры эмоций в виде ограниченного участка нервной ткани. Каждый, кто пытается рассмотреть организацию церебрального субстрата эмоций, непременно говорит о системе, о широко разветвленной констелляции нервных образований, представленных на различных уровнях головного мозга высших животных и человека.
Однако системный подход продуктивен только в том случае, если мы более или менее определенно указываем на конкретные элементы, из которых состоит данная система, на функциональное значение каждого из этих элементов и на правила взаимодействия элементов друг с другом при функционировании системы в целом. Причем под элементами мы подразумеваем не гипотетические «блоки» логических схем, а реальные мозговые образования, ту приуроченность динамики к структуре, которую И. П. Павлов считал фундаментальным принципом своей материалистической теории. К сожалению, именно последнее требование оказалось критическим моментом для нейрофизиологического обоснования целого ряда очень красивых и логически стройных концепций. Иными словами, любая концепция, претендующая на объяснение принципов организации поведения, должна быть сопоставима с анатомическим строением мозга, поскольку и первое, и второе формировались в русле единого эволюционного процесса.
Наш собственный подход к этой проблеме базируется на потребностно-информационной теории эмоций. В обычных, неосложненных условиях мозг ориентирует поведение на высоковероятные события — на этом принципе строится адекватное приспособление к среде. При выработке переключения разнородных условных рефлексов, по Э. А. Асратяну [2], когда один и тот же сигнал (звонок) утром подкреплялся пищей, а вечером — болевым электрораздражением, реагировать целесообразно — значит утром бежать к кормушке, а вечером спасаться от тока в безопасной половине камеры. В отличие от обезьян и собак выработка переключения представляет трудную задачу для крыс. Мы были немало удивлены, когда М. Л. Пигарева [10] показала, что двустороннее разрушение гиппокампа резко облегчает формирование этой формы поведения у крыс. Мы предположили, что поведение крыс с поврежденным гиппокампом избирательно ориентировано на сигналы высоковероятных событий, например, на получение пищи в пищевой обстановке. При этом поведение перестает осложняться прогнозом событий, маловероятных в данной ситуации, в опытах Пигаревой — возможностью болевого раздражения в пищевой обстановке.
Справедливость сделанного вывода была подтверждена опытами Пигаревой с частичным случайным подкреплением пищевых условных рефлексов (рис. 2). При случайном подкреплении 50 % предъявленных условных сигналов выработка рефлексов у гиппо-кампэктомированных крыс существенно не отличается от аналогичной процедуры у интактных животных. Когда
51
же процент подкреплений снижается до 33—25, гиппокампэктомированные животные обнаруживают свою неспособность к образованию условных рефлексов на протяжении 10 дней опыта. Интактные крысы достигают высоких уровней реагирования к 8—9-му дню.
Итак, если в мозгу хранятся условные связи одного и того же стимула со следами двух событий, ранее следовавших за этим стимулом, то в отсутствии гиппокампа будет активирована условная связь сигнала с высоковероятным подкреплением. По данным А. Я. Мехедовой [5], особое значение для оценки вероятности подкрепления у собак имеет латеральная область префронтального неокортекса. Что касается гиппокампа, то он обеспечивает реакции на сигналы маловероятных событий.
В каких случаях целесообразно такое реагирование? В случае дефицита информации, необходимой для организации эффективного поведения, т. е. в случае, когда в процесс организации поведения вовлекается нервный механизм эмоций. По-видимому, гиппокамп участвует в сравнении наличного стимула с энграммами, хранящимися в памяти не только в процессе ввода информации в долговременную память, но и в процессе извлечения следов из памяти для их использования в текущем поведении.
Если переход от избирательного реагирования на строго определенные сигналы к генерализованным реакциям на все более широкий круг стимулов зависит от степени эмоционального напряжения, то какую роль здесь играет второй фактор, детерминирующий эмоцию,— сила актуализированной потребности? Интересные факты на этот счет М. Л. Пигарева [10] получила в опытах с выработкой переключения у интактных и амигдалэктомированных крыс (табл. 2).
Табл. 2 наглядно показывает, что после двустороннего повреждения ядер миндалевидного комплекса выработка условнорефлекторного переключения у крыс возможна только при сочетании слабого болевого раздражения с высокой пищевой возбудимостью или при использовании сильного тока после короткой пищевой депривации.
Таблица 2
ЗАВИСИМОСТЬ ВЫРАБОТКИ УСЛОВНОРЕФЛЕКТОРНОГО ПЕРЕКЛЮЧЕНИЯ ОТ СИЛЫ ТОКА И ПРОДОЛЖИТЕЛЬНОСТИ ГОЛОДАНИЯ
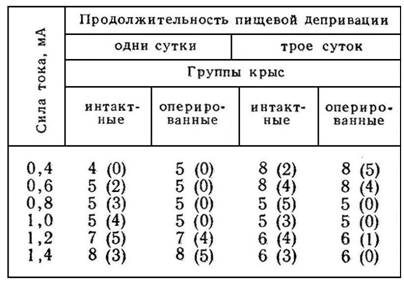
Примечание. Цифры показывают число крыс в каждой группе, цифры в скобках — число крыс, выработавших переключение (три опыта подряд со 100%-ным осуществлением как пищевого, так и оборонительного условных рефлексов) на протяжении 60 дней.
Иными словами, в пищевой ситуации, когда условный сигнал активирует обе связи — высоковероятную с пищей и маловероятную с болью — пищевая реакция у амигдалэктомированных крыс осуществится и подавит потенциально возможную оборонительную только при явном преобладании голода над потребностью избегания боли. По-видимому, миндалина необходима для тонкой оценки конкурирующих потребностей, для обеспечения их баланса, их динамического сосуществования в данной конкретной ситуации. Если разрушение гиппокампа превращает животное в автомат, реагирующий только на сигналы высоковероятных событий и игнорирующий все другие альтернативы, то повреждение миндалины однозначно ориентирует поведение на удовлетворение доминирующей потребности без учета других мотиваций. В естественной среде обитания такое поведение может оказаться крайне нецелесообразным: например, мотивированное голодом животное будет пренебрегать сигналами опасности.
Данные М. Л. Пигаревой хорошо согласуются с результатами наших опытов с выработкой условной реакции избегания у крыс при болевом
52
раздражении другой особи. Оказалось, что последствия амигдалэктомии зависят от того, какая из двух конкурирующих мотиваций относительно преобладала до операции у данного животного: чувствительность к крику боли другой крысы или предпочтение закрытой со всех сторон педали (включавшей ток) открытому пространству экспериментальной камеры.
Сходные факты получили Уайт и Вейнгартен [25]. Сытые амигдалэктомированные крысы проявляли большую исследовательскую активность, чем контрольные, в то время как пищевая активность крыс в обеих группах была одинаковой. Исследовательское поведение голодных амигдалэктомированных крыс уступало аналогичному поведению контрольных, тогда как пищевая активность оперированных животных преобладала над активностью контрольных. Таким образом, в опытах Уайта и Вейнгартена амигдалэктомия усиливала то поведение, которое инициировалось доминирующей потребностью. Справедливое утверждение Глура [20] о принадлежности миндалины к системе структур, определяющих выбор поведения, можно принять с уточнением, что миндалина участвует в этом выборе путем «взвешивания» конкурирующих эмоций, порожденных конкурирующими потребностями.
О конкуренции именно эмоций, а не самих потребностей (мотиваций) свидетельствуют следующие факты. Показано, что базолатеральная часть миндалины связана с влиянием прошлого опыта утоления жажды, а не с «тканевой жаждой», не с детекцией водно-солевого баланса. Повреждение миндалин влияет на реакции, вызванные у кошек страхом, а не болью. Вот почему повреждение миндалин у собак нарушает классические условные оборонительные рефлексы и не сказывается на инструментальных, где признаки страха исчезают по мере совершенствования защитной условной реакции. Все сказанное выше позволяет предполагать, что миндалина вовлекается в процесс организации поведения на сравнительно поздних этапах этого процесса, когда актуализированные потребности уже сопоставлены с перспективой их удовлетворения и трансформированы в соответствующие эмоциональные состояния. Миндалина принадлежит к системе тех мозговых образований, которые реализуют «прерывающую функцию» эмоций, ориентируя поведение на первоочередное удовлетворение главенствующей потребности с учетом наличной ситуации и прошлого опыта животного.
Повреждения гипоталамуса зачастую дают эффект, противоположный амигдалэктомии: животные перестают реагировать на «тканевую жажду» и падение содержания глюкозы в крови, но продолжают отвечать на условные сигналы воды и пищи. Их прошлый опыт приобретает известную самостоятельность, будучи изолированным от текущих нужд организма. В опытах с реакцией избегания у крыс при болевом раздражении другой особи мы наблюдали результат, противоположный последствиям амигдалэктомии. Если амигдалэктомия нарушает баланс между конкурирующими мотивациями и выявляет доминирующую из них, то двустороннее повреждение гипоталамуса, напротив, выравнивает силу мотиваций. Для этих крыс характерно «застревание» между педалью, включавшей ток, и открытым пространством камеры.
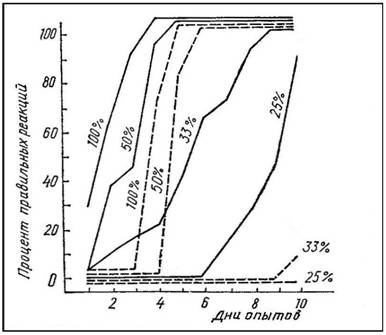
Рис. 2. Процент правильных реакций при выработке условных двигательных пищевых рефлексов у контрольных (сплошные линии) и гиппокампэктомированных (прерывистые линии) крыс.
Цифры на графиках обозначают процент случайно подкрепляемых предъявлений условного сигнала.
По оси абсцисс — дни опытов
53
Всю совокупность экспериментальных данных о роли неокортекса, гиппокампа, миндалины и гипоталамуса в генезе эмоциональных состояний и, соответственно, в организации целенаправленного поведения мы попытались обобщить схемой, представленной на рис. 5. Разумеется, эта схема огрубляет и упрощает реальную сложность взаимодействия мозговых структур, игнорирует многие другие функции этих мозговых образований. Вместе с тем мы убеждены в продуктивности именно простых схем, потому что только они доступны прямой экспериментальной проверке.
III. МОЗГ И ИНДИВИДУАЛЬНЫЕ ОСОБЕННОСТИ ПОВЕДЕНИЯ
Основывая свою классификацию типов высшей нервной деятельности на параметрах силы, подвижности и уравновешенности процессов возбуждения и торможения, И. П. Павлов одновременно указывал на значение индивидуальных особенностей взаимодействия макроструктур головного мозга. Так, он связывал «мыслительный тип» человека с функциональным доминированием лобных отделов неокортекса, а «художественный тип» — с преобладанием других отделов новой коры и «эмотивных» подкорковых образований. Развивая идеи Павлова, Б. М. Теплов и В. Д. Небылицын выделили общие свойства нервной системы: активность и эмоциональность. В. Д. Небылицын [6] предположил, что активность зависит от индивидуальных особенностей функционирования системы: передние отделы новой коры — активирующая ретикулярная формация мозгового ствола, а эмоциональность определяется взаимодействием фронтального неокортекса с лимбической системой мозга. Согласно Г. Айзенку [19], в основе параметра силы нервной системы, равно как и степени экстра-интроверсии, лежит взаимодействие восходящей активирующей системы с передними отделами новой коры. Модифицируя схему Айзенка, Д. Грей [21] добавил к этим двум структурам гиппокамп и медиальную часть перегородки.
Показано, что разрушение одних и тех же лимбических структур у животных разных генетических линий ведет к существенно различным последствиям. С другой стороны, мыши, принадлежащие к линиям с различным объемом гиппокампа и неокортекса, обладают характерными особенностями поведения в тесте открытого поля и при выработке условной реакции избегания.
Исследования нашей лаборатории привели к выводу о ведущей роли четырех мозговых структур в генезе эмоциональных реакций и в организации целенаправленного поведения. Мы имеем в виду передние отделы новой коры, гиппокамп, миндалину и гипоталамус. Специализация функций перечисленных мозговых структур дает основание для предположения о том, что индивидуальные особенности активности каждой из них и тем более особенности их взаимодействия во многом определяют индивидуальные (типологические) черты поведения животных. Ниже излагаются результаты двух серий экспериментов, подкрепляющих, по нашему мнению, высказанное предположение.
Первый эксперимент. Л. А. Преображенская [12] систематически исследовала электрическую активность гиппокампа при выработке условнорефлекторного переключения у собак. У 8 собак сначала вырабатывали инструментальный пищевой рефлекс надавливания правой передней лапой на педаль в ответ на звуковой условный сигнал (тон). Затем тот же условный сигнал, подаваемый на фоне действия сигнала переключателя (шум и мелькание лопастей вентилятора), начинали подкреплять болевым раздражением лапы электрическим током. Это раздражение собака могла прервать или полностью предотвратить подъемом левой передней лапы до определенного уровня. Детали выработки пищевых и оборонительных инструментальных условных рефлексов описаны ранее [11].
Оказалось, что при переходе от пищевой ситуации к оборонительной происходит учащение гиппокампального тета-ритма у всех собак. Вместе с тем каждое животное характеризуется своим
54
диапазоном изменений частотного спектра регулярной активности ЭГГ, и этот диапазон коррелирует с динамикой выработки переключения условных рефлексов. У собак с более частым тета-ритмом выработка переключения произошла сравнительно быстро и легко: они стали реагировать на условный сигнал в соответствии с наличной ситуацией после 5—6 опытов. Иная картина наблюдалась у собак, условнорефлекторная активность которых носила неустойчивый, волнообразный характер с тенденцией к невротизации. Аналогичные данные были получены и в опытах с другими собаками. Животные с относительно медленным гиппокампальным тета-ритмом отличались малой общительностью, безразличным отношением к экспериментатору. Они испытывали затруднения и при решении других задач, связанных с изменением рода деятельности.
Полученные факты совпадают с имеющимися в литературе данными о корреляции доминирующей частоты гиппокампального тета-ритма у крысе уровнем исследовательской активности, характерным для каждого животного. Оба эти показателя достаточно стабильны у одной и той же крысы. Таким образом, можно сказать, что индивидуально типичный для данного животного диапазон изменений частоты гиппокампального тета-ритма отражает параметр, который Павлов обозначил как инертность (или, напротив, подвижность) нервной системы. Если учесть ту роль, которую, согласно современным представлениям, играют механизмы возвратного торможения в генезе ритмических колебаний биопотенциалов, то положение Павлова о подвижности нервных процессов возбуждения и торможения наполняется конкретным нейрофизиологическим содержанием. С другой стороны, данные об участии гиппокампа в процессах регистрации информации, равно как и в процессах ее извлечения из памяти делают понятной связь индивидуальных особенностей гиппокампального тета-ритма с индивидуальными особенностями поведения. Ранее мы показали, что непосредственно перед осуществлением инструментального условного рефлекса возрастает степень пространственной синхронизации электрической активности гиппокампа и гипоталамуса, что свидетельствует об усилении функционального взаимодействия этих двух структур на данном этапе целенаправленного поведения [9].
Второй эксперимент. Исследование реакций избегания, подкрепляемых включением искусственных стимулов (усиление освещенности и звучание тона) или биологически специфическим воздействием (крик боли партнера), показало, что для большинства крыс (около 60%) более эффективным стимулом является крик боли. Примерно 30% животных сильнее реагируют на включение света и звука, а 10 % одинаково высокочувствительны к обоим разновидностям аверсивных стимулов. Трудно сказать, в какой мере эти индивидуальные особенности поведения зависят от генетических или средовых факторов, хотя имеются сведения о том, что частота нажатий на рычаг, подкрепляемых включением света и ослаблением шума, на 71% детерминирована у лабораторных крыс генетически. Представляло интерес выяснить, в какой мере эти особенности поведения зависят от сохранности лимбических структур.
Опыты поставлены на 20 беспородных белых крысах-самцах. Экспериментальная камера представляла деревянный ящик размерами 33×41×34 см. Внутри ящика имелись: 1) относительно просторная часть площадью 33×23 см; 2) плексигласовый «домик» площадью 16×14 см с постоянно открытой дверью и полом-педалью, давление на которую автоматически включало счетчик времени; 3) расположенное рядом с «домиком» за тонкой прозрачной звукопроницаемой перегородкой помещение для партнера с полом в виде металлической решетки. Вся камера освещалась рассеянным светом от лампы мощностью 100 Вт, установленной под потолком.
Исследуемое животное ежедневно помещали в большое отделение камеры на 5 мин и регистрировали время его пребывания в «домике» на педали. В течение первых 5 дней каждое появление крысы в «домике» приводило
55
к включению дополнительного освещения лампой 100 Вт, расположенной в 45 см от пола камеры, и звукового стимула — тона частотой 220 Гц и громкостью 80 дБ. На протяжении следующих 5 дней вход в «домик» сопровождался электрическим раздражением лап крысы-«жертвы» силой 1—2 мА. Раздражение «жертвы» продолжалось 3—5 с с 5-секундными интервалами до тех пор, пока исследуемая крыса находилась на педали. Последние 5 дней вход в «домик» снова усиливал освещение и включал звук.
На рис. 3 график 1 показывает среднее время пребывания на педали 7 интактных крыс, для которых сигналы оборонительного возбуждения партнера (крик, движения, выделение специфических пахучих веществ) были более эффективным стимулом, чем усиление освещенности и звучание тона. После двусторонней коагуляции фронтальных отделов новой коры и гиппокампа у этих крыс наблюдались прямо противоположные отношения: время пребывания на педали при действии звука и света уменьшилось, а при крике «жертвы» возросло (график II на рис. 3); 5 крыс с двусторонним повреждением фронтальной коры, латерального и вентромедиального гипоталамуса оказались в равной мере чувствительными и к комбинации звука с усилением освещенности, и к сигналам оборонительного возбуждения партнера (рис. 4). Эти животные отличались пугливостью, повышенной
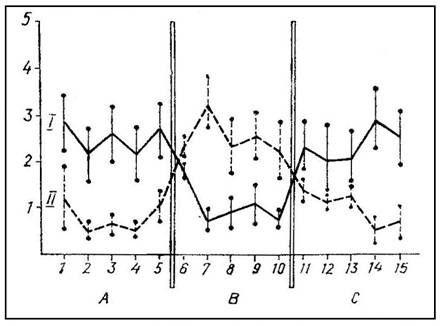
Рис. 3. Среднее время пребывания на педали при действии света и звука (А, С) или крика партнера (В) у интактных крыс (I) после двустороннего повреждения фронтальной коры и гиппокампа (II).
Абсцисса — дни опытов, ордината — время в мин.
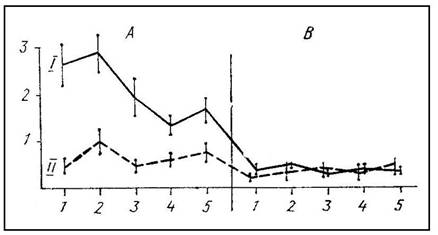
Рис. 4. Среднее время пребывания на педали при действии света и звука (I) или крика партнера (II) до (А) и после (В) двустороннего повреждения фронтальной коры и гипоталамуса.
Остальные обозначения, как на рис. 3
агрессивностью, бурными реакциями на прикосновение наряду с признаками ослабления аверсивности открытого пространства. Крысы медленно и редко заходили в «домик», а при включении света и звука или при крике партнера выходили из «домика» спустя 10—20 с. Если что-либо отвлекало крысу (например, она начинала чистку шерсти), свет, звук и крик «жертвы» теряли свою эффективность.
Мы полагаем, что результаты этих опытов согласуются с фактами, полученными ранее при раздельном разрушении у крыс фронтального неокортекса, гиппокампа, миндалины и гипоталамуса. Одновременное повреждение структур «информационной системы» (фронтальный неокортекс и гиппокамп) делает крыс высокочувствительными к ранее малоэффективным искусственным стимулам (свет и звук) и в то же время снижает их реактивность по отношению к зоосоциальным сигналам о состоянии другой особи того же вида. Что касается повреждения фронтальной коры, латерального и вентромедиального гипоталамуса, то усиленная невротическая реакция крыс на любой внешний стимул сочетается у этих животных с неспособностью избирательно реагировать на сигналы различного биологического значения.
Оценивая всю совокупность имеющихся в настоящее время фактов, мы склонны высказывать предположение о том, что индивидуальные особенности соотношения «информационной системы» фронтальная кора — гиппокамп с «мотивационной системой» миндалина
56
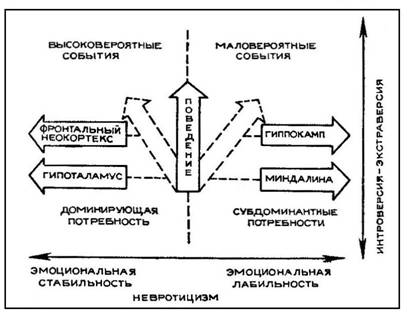
Рис. 5. Гипотетическая схема зависимости параметров интро-экстраверсии и невротицизма от индивидуальных особенностей взаимодействия четырех мозговых структур
— гипоталамус лежат в основе параметра экстра-интроверсии (рис. 5). Соотношение систем фронтальная кора — гипоталамус и миндалина — гиппокамп определяет другой параметр индивидуальных особенностей поведения, близкий по своим характеристикам параметру невротизма или эмоциональности. С этой точки зрения, павловская шкала силы или слабости нервной системы больше соответствует шкале невротизма, а не экстра-интроверсии, как полагает Г. Айзенк [19]. Для фактора подвижности или инертности, в павловском его понимании, большое значение имеет активность системы гипоталамус — гиппокамп и ее соотношение с системой фронтальная кора — ядра миндалевидного комплекса. Наше общее предположение сводится к тому, что индивидуальные особенности взаимодействия этих четырех мозговых структур лежат в основе индивидуальных особенностей поведения описанных ранее античными авторами в виде темпераментов, типов высшей нервной деятельности Павлова и шкалы экстра-интроверсии. Различные формы нарушения нормального взаимодействия перечисленных структур, возможно, определяют основные разновидности неврозов.
Полвека назад Л. С. Выготский [4; 127] писал, что «психология чувств оказалась самой бесплодной и скучной из всех глав этой науки». Потребностно-информационный подход открывает новые перспективы в исследовании нейрофизиологии, анатомии и психофизиологии эмоций.
1. Анохин П. К. Эмоции. — Большая медицинская энциклопедия, 2-е изд., 1964, т 35, с. 339—357.
2. Асратян Э. А. Лекции по некоторым вопросам нейрофизиологии. — М., 1959.
3. Асратян Э. А. Рефлекторная теория и проблема мотиваций. — В кн.: Основные проблемы электрофизиологии головного мозга. — М.. 1974, с. 5—20.
4. Выготский Л. С. Спиноза и его учение об эмоциях в свете современной психоневрологии. — Вопросы философии, 1970, № 6, с. 119—127.
5. Мехедова А. Я. О роли лобных областей мозга в формировании условных реакций, адекватных величине и вероятности их подкрепления. — Журнал высшей нервной деятельности, 1971, т. 21, № 3, с. 459—464.
6. Небылицын В. Д. К вопросу об общих и частных свойствах нервной системы. — Вопросы психологии, 1968, № 4, с. 29—43.
7. Осипов В. П. О физиологическом происхождении эмоции. — В кн.: Сборник, посвященный 75-летию акад. И. П. Павлова. Л., 1924, с. 109—110.
8. Павлов И. П. Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных. — М., 1973.
9. Павлыгша Р. А., Труш В. Д., Михайлова Н. Г., Симонов П. В. Взаимоотношение электрической активности мозговых структур в процессе мотивированного поведения. — В кн.: Функциональная значимость электрических процессов головного мозга. М., 1977, с. 58—67.
10. Пигарева М. Л. Лимбические механизмы переключения (гиппокамп и миндалина). — М., 1978.
11. Преображенская Л. А. Эмоциональное напряжение при условнорефлекторном переключении инструментальных разнородных рефлексов. — Журнал высшей нервной деятельности, 1978, т. 28, № 3, с. 505—513.
12. Преображенская Л. А. Индивидуальные особенности гиппокампальной электрической активности и условнорефлекторной деятельности у собак. — Журнал высшей нервной деятельности, 1981, т. 31, № 3, с. 479—488.
13. Симонов П. В. О соотношении двигательного и вегетативного компонентов условного оборонительного рефлекса у человека. — В кн.: Центральные и периферические механизмы двигательной деятельности животных и человека. М., 1964, с. 65—66.
14. Симонов П. В. Двигательный стереотип и физиология эмоций. — В кн.: XVIII Международный психологический конгресс. Симпозиум 3. М., 1966, с. 97—99.
15. Симонов П. В. Что такое эмоция? — М., 1966.
16. Симонов П. В. Эмоциональный мозг.— М., 1981.
17. Ухтомский А. А. Собрание сочинений, т. 1. —Л., 1950.
18. Ухтомский А. А. Доминанта. — М.; Л.. 1966.
Поступила в редакцию 2.VII.1982 г.