116
ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ
ФАНТОМНЫЕ ОБРАЗЫ БИНОКУЛЯРНОГО ЗРЕНИЯ
В СИСТЕМЕ МЕЖПОЛУШАРНЫХ ОТНОШЕНИЙ
В. В. СУВОРОВА, М. А. МАТОВА
Предметом настоящего исследования является бинокулярное зрение в системе межполушарных отношений как функция анализатора, имеющего парный рецептор и парное мозговое управление. Данная работа, выполненная в плане широкого комплексного исследования функциональной асимметрии в работе больших полушарий головного мозга, отражает круг вопросов, связанных с особенностями формирования зрительных образов. Наше внимание привлекли некоторые феномены бинокулярного зрения, которые вполне объяснимы, если рассматривать глаз только как оптическую систему, но которые еще мало понятны по своим психофизическим механизмам.
К таким феноменам можно отнести, в частности, фузию, т. е. слияние двух монокулярных изображений в единый образ. В рамках физиологической оптики фузия рассматривается как результат корреспонденции сетчаток двух глаз. Однако совершенно очевидно, что образование единого образа в анализаторе, имеющем два рецептора и два мозговых центра, должно опосредствоваться определенными закономерностями билатерального взаимодействия и латерализации церебральных функций [1], [8] и др.
Известно, что в центральном бинокулярном зрении корреспондируют центральные ямки сетчаток (фовеа), а в периферическом — носовая зона сетчатки одного глаза и височная — другого. Нас интересовало, какую роль играет каждая из этих зон в образовании фузионного образа и обладают ли зоны одинаковыми функциями. Открытия последних лет в области асимметрии межполушарных отношений дают основания для предположения, что фузия как продукт межполушарного взаимодействия является суммой не равных слагаемых. Гипотезу о различии функций отдельных зон сетчаток глаз в формировании фузионного образа можно обосновать следующими анатомо-физиологическими данными:
1. Носовые и височные зоны центрального и периферического зрения (за исключением фовеальных зон) по-разному взаимосвязаны с мозговыми полушариями. От носовых зон нервные пути идут к противоположным (контралатеральным) полушариям, перекрещиваясь в хиазме, а от височных зон они идут без перекреста к одноименным (ипсилатеральным) полушариям [15], [17], [32], [33].
2. Морфологическое строение носовых и височных зон периферического зрения не вполне идентично: в носовых зонах колбочек больше, чем в височных, особенно на периферии [14], [17], [28].
3. Волокна от носовых и от височных зон периферического и центрального (желтого пятна) зрения приходят в разные подслои IV слоя зрительной коры 17-го поля, имеющие разное морфологическое строение (нервные клетки разной формы, разного размера и. разной густоты расположения) [28], [29], [32], [33] и др.
4. Рецептивные поля бинокулярных нейронов, расположенные в носовых зонах, имеют более точную топографическую проекцию в зрительную кору по сравнению с рецепторами височных зон, что доказано исследованиями на нейронном уровне [34], [36] и др.
5. Участие носовых и височных зон периферического зрения в построении фузионного образа не одинаково. В условиях бинокулярного зрения височные зоны целиком участвуют в построении бинокулярного поля зрения, а носовые зоны имеют участки (крайней периферии), которые функционируют как исключительно монокулярные (зоны височного полулуния) [14], [28] и др.
Все эти различия могут иметь и, вероятнее всего, имеют определенное значение в билатеральной регуляции фузионного процесса. Для того чтобы сравнить функции корреспондирующих зон в образовании фузионного образа в центральном и периферическом зрении, необходимо было расчленить каждую такую пару корреспондирующих зон, заставить их работать самостоятельно и независимо. Проще говоря, требовалось создать условия изолированной проекции раздражителей в рецепторы каждой из зон центрального и периферического зрения.
МЕТОДИКА И УСЛОВИЯ ПРОВЕДЕНИЯ ЭКСПЕРИМЕНТА
Задача расчленения функций корреспондирующих зон сетчаток обоих глаз методически очень проста и не требует сложной аппаратуры. Опыты проводились на приборе, главной деталью которого был маленький экран (размером 45×100 мм), располагаемый у глаз испытуемого. Этот экран, сделанный из матового полупрозрачного стекла, был заключен в пластмассовый кожух, ограничивающий поле зрения. Внутри прибора находилась съемная перегородка, разделяющая его на два изолированных бокса с одинаковыми экранами: правым — для правого глаза и левым — для левого. Оба экрана были снабжены измерительными шкалами с началом отсчета от разделительной перегородки: левый экран с отрицательными значениями, правый — с положительными. Вдоль экрана проходила прорезь, куда вставлялись слайды со стимульным материалом. Расстояние от слайдов до глаз испытуемых составляло в среднем
117
65 мм. Разделительная перегородка обеспечивала гаплоскопическое (раздельное) зрение испытуемых: каждый глаз видел только один, свой, экран.
Все опыты проводились в условиях первичного положения глаз (фиксационные точки расположены в центре каждого экрана, зрительные оси параллельны), которое достигалось путем предварительной тренировки испытуемых. Индикатором соблюдения требования о первичном положении глаз было слияние фиксационных точек. Такое положение глаз позволяло довольно точно проецировать раздражители в заданные зоны сетчаток. Отсутствие задач, адресованных к остроте зрения, обусловливало достаточно четкое видение раздражителей для определения локализации их в зрительном поле (главная задача испытуемых).
По команде экспериментатора испытуемому следовало поднести прибор к глазам и, глядя на экран, сначала описать местоположение предъявляемых стимулов словесно, а затем показать с внешней стороны экрана указкой, где он видит стимулы на просвет. Время предъявления раздражителей не лимитировалось и зависело от темпа сенсомоторных действий испытуемого. Речевые и моторные ответы испытуемых регистрировались экспериментатором в специальных бланках протоколов.
Следует особо отметить, что в этих экспериментах мы создавали для глаз испытуемого как оптической системы необычные для естественного бинокулярного зрения условия, помещая раздражители ближе зоны ясного видения и исключая конвергенцию. Однако с позиций билатеральной регуляции фузионных процессов восприятие этой области пространства, обычно не анализируемое, представляет особый интерес. Если принять, что «возбудителем зрительного ощущения является пространственная и временная конфигурация лучистой энергии» [9; 377], то в поле зрения нет и не должно быть «мертвой зоны». Следовательно, постановку наших опытов с методической стороны нужно признать правомерной: стимул, предъявляемый из любой точки пространства и проецируемый в зрительные рецепторы, должен вызывать и вызывает соответствующую зрительную перцепцию.
Опыты проведены со взрослыми испытуемыми, которые не предъявляли жалоб на недостаточность бинокулярного зрения. В данных экспериментах принимали участие 50 человек (выборка условно принята нами за норму в отношении латерализации мозговых функций без диагностированного левшества). Кроме того, были обследованы лица с отклонениями от нормального распределения функций: а) с симметричным распределением речевых функций (больные логоневрозом); б) с симметричной и зеркальной латерализацией моторных функций (амбидекстрией и левшеством) — монозиготные и дизиготные близнецы.
С каждым испытуемым проводилось 8 серий опытов (по 6 предъявлений в каждой серии). С некоторыми — опыты многократно повторены через разное время. Результаты, полученные при этом, оказались одинаковыми или почти одинаковыми. В настоящем сообщении в соответствии с выделенной проблемой будут представлены данные только трех серий опытов и только первой выборки испытуемых.
РЕЗУЛЬТАТЫ ЭКСПЕРИМЕНТОВ И ИХ ОБСУЖДЕНИЕ
В I серии опытов, поставленных для изучения роли корреспондирующих зон периферического зрения в образовании фузионного образа, стимулы проецировались из наружных частей монокулярных полей зрения в носовые зоны сетчаток левого и правого глаза. Зрительные стимулы (две вертикальные линии, одинаковые по размеру, но разные по цвету) помещались симметрично от разделителя полей зрения — на отметках «—40» для левого глаза и «+40» для правого глаза.
В результате опытов обнаружено, что 90 % испытуемых видели и показывали стимулы, проецировавшиеся в носовые зоны сетчатки, в соответствии с их действительным местоположением. Испытуемые видели единое поле зрения (результат совмещения монокулярных полей), в центре которого находилась точка (фузионный образ двух фиксационных точек). В дальнейшем эту точку мы будем условно называть фузионной. Вдоль наружных границ поля зрения испытуемые видели предъявляемые стимулы: правый — справа и левый — слева. Они безошибочно показывали на внешней стороне экрана местоположение этих стимулов: левый показывали на отметке «—40» левого экрана, а правый — соответственно на отметке «+40» правого экрана (рис. 1, А)
Таким образом, по результатам I серии экспериментов можно констатировать адекватное восприятие пространственного положения стимулов и их непосредственную чувственную перцепцию, т. е. формирование их перцептивного образа.
Во II серии опытов стимулы проецировались из внутренних частей монокулярных полей зрения в височные зоны сетчаток левого и правого глаза: те же линии помещались слева и справа от разделителя полей зрения — соответственно на отметках «—10» и «+10» левого и правого экранов.
В итоге этой серии опытов было установлено, что в 80°/о случаев при одновременной проекции двух раздражителей в височные зоны сетчаток испытуемые видели единое поле зрения, в центре которого находилась фузионная точка, а на крайней периферии, вдоль наружных границ поля зрения, симметрично виделись раздражители. И зрительно, и моторно эти раздражители локализовались на экране в точках, не соответствующих их действительному положению. Раздражитель, предъявляемый правому глазу, который находился на отметке «+10» правого экрана, испытуемые видели у левой границы зрительного поля и в соответствии с этим показывали указкой на отметках «—40», «—45». И, наоборот, раздражитель, предъявляемый левому глазу, находившийся на отметке «—10», они видели у правой границы зрительного поля и показывали в зоне отметок «+40»,«+45» (рис. 1, Б). В остальных 20 % случаев испытуемые видели и локализовали двигательно один из стимулов или оба в тех же точках пространства, где они находились в действительности.
Анализ экспериментальных данных показывает, что в тех 80% случаев, когда испытуемые не видели стимул в точке его действительного местонахождения, т. е. в пространстве,
118
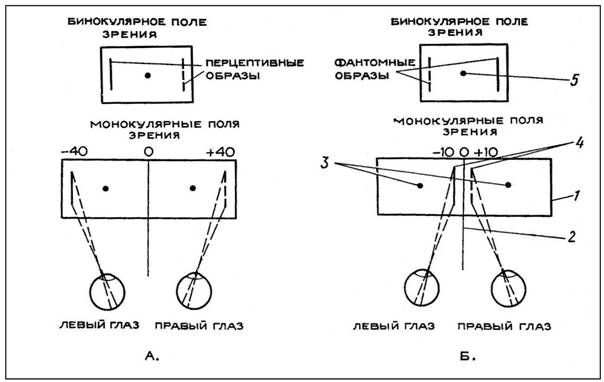
Рис. 1, А и Б. Схема проекции образов в бинокулярном поле зрения. А. При проекции стимулов в носовые зоны сетчаток. Б. При проекции стимулов в височные зоны сетчаток. Условные обозначения: 1 — экран; 2 — перегородка; 3 — фиксационные точки; 4 — стимулы; 5 — фузионная точка.
которое проецируется в височную зону сетчатки1 (и в котором стимул в действительности находился), они видели его и в соответствии с этим моторно локализовали смещенным в контралатеральном направлении, т. е. в пространстве, «принадлежащем» носовой зоне другого глаза. Таким образом, стимул, предъявляемый через височную зону сетчатки одного глаза, в большинстве случаев воспринимался как расположенный в пространстве корреспондирующей с ней (носовой) зоны другого глаза.
Подобное смещение локализации стимула в контралатеральном направлении наблюдалось и у оставшихся 10% испытуемых, данные которых не вписались в общую закономерность результатов I серии опытов. Эти испытуемые воспринимали пространственное положение стимула, проецируемого в носовую зону сетчатки, тоже не адекватно: они видели и показывали его указкой в пространстве не своей, а корреспондирующей (височной) зоны другого глаза.
Таким образом, в абсолютном большинстве случаев при проекции раздражителей в височные зоны сетчаток обоих глаз и в незначительном количестве случаев при проекции раздражителей в носовые зоны стимулы воспринимались в точках пространства, не соответствующих их действительному местоположению. В силу каких-либо причин перцептивные образы предъявляемых раздражителей не создавались. Вместо них возникали зрительные образы, которые по отношению к раздражителям виделись смещенными в контралатеральном направлении, и которые мы назвали фантомными.
Постепенно стало очевидно, что фантомные образы существенно отличаются от перцептивных. Так было установлено, что фантомные образы не устойчивы. Если в зоне локализации фантомного образа возникал движущийся объект (испытуемый совершал стирающие движения указкой с наружной стороны экрана), то фантомный образ исчезал на короткий промежуток времени и зрительное поле становилось пустым. При этом исчезновение фантомного образа никогда не приводило к появлению перцептивного образа, хотя стимул, порождавший фантомный образ, по-прежнему находился в поле зрения испытуемого. Испытуемый сообщал, что на мгновение объект наблюдения исчез, стерся, но сразу же снова появился, проступил на прежнем месте, либо чуть отодвинулся в сторону. Легкость, с которой образ исчезал, быстрота его восстановления и смещение в сторону обусловливались индивидуальными особенностями испытуемых.
Подводя итог опытам с бинокулярным периферическим зрением, надо отметить, что функции двух членов каждой из пар корреспондирующих зон сетчаток оказались, как мы и предполагали, не одинаковыми: через носовые зоны в периферическом зрении чаще формируется перцептивный образ, т. е. происходит адекватная
119
фиксация положения стимула в пространстве, а через височные зоны — чаще формируется фантомный образ. Причем в большинстве случаев фантомные образы локализуются в пространстве, принадлежащем носовым зонам.
Встает закономерный вопрос: а как образуется фузионный образ в центральном бинокулярном зрении, где корреспондируют центральные ямки сетчаток? Напомним, что центральная ямка желтого пятна (фовеа) по целому ряду данных, и в первую очередь клинических, имеет двойную проекцию в оба полушария. При разрушении зрительной коры одного полушария периферическое поле зрения выпадает, а центральное (фовеальное) полностью сохраняется. Если каждая фовеальная зона имеет двойную проекцию и с одним полушарием связана прямыми, а с другим перекрестными связями, то естественно допустить, что каждая из фовеальных зон может функционировать и как носовая (связанная с контралатеральным полушарием), и как височная (связанная с ипсилатеральным полушарием), т. е. что каждая фовеальная зона может и фиксировать объект в пространстве, создавая его перцептивный образ, и продуцировать фантомные образы.
В III серии опытов, где изучались особенности образования фузионного образа в центральном зрении, раздражителями служили только точки (диаметром 4 мм), которые располагались в центре каждого экрана перед фовеальной зоной каждого глаза. Периферическая зона сетчаток при этом не стимулировалась. В первой части опыта черная точка помещалась перед фовеальной зоной правого глаза, левый экран оставался в это время совершенно пустым.
В этих условиях испытуемые видели точку не справа, где она им предъявлялась, а в центре бинокулярного зрительного поля. На предложение экспериментатора показать видимую ими точку 70°/о испытуемых указали ее место в центре правого экрана. Остальные 30 % испытуемых хотя и видели точку тоже в середине бинокулярного поля зрения, но показывали ее не на правом экране, где она была в действительности, а на левом экране, где никакой точки на самом деле не было.
Таким образом, и в центральном зрении у части испытуемых наблюдалась неадекватная локализация стимула. Так же как в периферическом зрении образ стимула здесь смещался в контралатеральном направлении. Это смещение могло быть стойким: оно имело место при многократном повторении опыта и стабильно воспроизводилось в разные экспериментальные дни. Во второй части опыта та же точка помещалась в центре левого экрана перед фовеальной зоной левого глаза, в то время как правый экран оставался совершенно пустым.
Результаты этих опытов в точности соответствовали результатам предыдущей серии.' Только теперь стимул локализовался правильно в половине случаев, а в половине случаев испытуемые снова показывали пустое место в центре другого экрана.
Во всех случаях неадекватной пространственной локализации раздражителя в центральном бинокулярном зрении вместо перцептивных образов, так же как в периферическом, формировались фантомные образы, которые обладали тем же свойством неустойчивости к внешнему воздействию. Фантомные образы центрального зрения, как и периферического, тоже временно исчезали при движении указки в местах их появления.
Местоположение фантомных образов в центральном зрении также не было случайным — проекция раздражителя в фовеальную зону одного глаза вызывала появление фантомного образа перед фовеальной зоной другого глаза, т. е. и в центральном зрении стимуляция одной из корреспондирующих зон могла привести к возникновению фантомного образа перед другой корреспондирующей зоной.
Итак, установлено, что фантомные образы возникают и в периферическом, и в центральном зрении; в периферическом зрении они чаще всего продуцируются височными зонами и передаются в пространство корреспондирующих с ними носовых зон; в центральном зрении фантомные образы появляются перед фовеальной зоной одного (чаще правого) глаза в ответ на предъявление раздражителя фовеальной зоне другого (чаще левого) глаза.
Анализ экспериментальных данных позволил нам выделить и описать основные свойства фантомных образов, которые коротко сводятся к следующему:
1. Фантомный образ возникает при непосредственном восприятии предъявляемого стимула и является его точным отображением.
2. Фантомному образу присуще определенное пространственное положение: в ответ на стимуляцию одной из корреспондирующих зон он возникает в пространстве другой корреспондирующей зоны, смещаясь в контралатеральном направлении. Местоположение такого образа в пространстве может задаваться преднамеренно, путем изменения местоположения стимула.
3. Фантомный образ характеризуется неустойчивостью к внешнему воздействию — он временно исчезает при движении дополнительного объекта в занимаемом им пространстве. Мера этой неустойчивости зависит от индивидуальных особенностей наблюдателя.
По всей вероятности, наличие фантомных образов в центральном и периферическом зрении нельзя объяснить с позиций геометрии бинокулярного зрения. Для доказательства этого предположения сравним данные I и II серий экспериментов.
Если исходить из понимания корреспонденции периферических зон сетчаток как соответствия точек, «которые лежат в одном и том же направлении и на одном и том же расстоянии от «fovea centralis» [17], то следует признать экспериментальные ситуации I и II серий наших опытов однотипными (Рис. I, А и Б). И в том и в другом случае происходит фузия фиксационных точек: наблюдатель видит две фиксационные точки как одну. Это свидетельствует о том, что и в том, и в другом варианте опытов имеет место наложение монокулярных, зрительных полей и образование бинокулярного поля зрения. Но если корреспондируют фовеальные зоны обоих глаз и две раздельно предъявляемые фиксационные точки сливаются и воспринимаются как одна, то должны корреспондировать и периферические зоны глаз. А если так, то раздражитель, предъявляемый височной зоне одного глаза может рассматриваться как находящийся в пространстве носовой
120
зоны другого глаза. Аналогично этому раздражитель, проецируемый в носовую зону одного глаза, может рассматриваться как находящийся в пространстве височной зоны другого глаза.
Однако в наших экспериментах такое наложение монокулярных полей зрения, в результате которого пространство корреспондирующих периферических зон оказывалось совмещенным, и раздражитель, предъявляемый одной из корреспондирующих зон, оказывался в пространстве другой корреспондирующей зоны, имело место только при проекции раздражителей в височные зоны сетчаток. И в этом случае перцептивные образы раздражителей заменялись фантомными образами.
Проекция раздражителей в носовые зоны подобного эффекта не вызывала, и в бинокулярном поле зрения раздражитель, проецируемый в носовую зону, воспринимался в пределах пространства той же зоны. Видимо, это можно объяснить закономерностями отношений латерального пространства с рецепторами зрительного анализатора: правое поле зрения проецируется в носовую половину сетчатки правого же глаза и левое — левого [22; 243]. Именно поэтому объект, предъявляемый монокулярно в носовую зону правого глаза, и в условиях наших экспериментов в бинокулярном поле зрения оставался справа. В этом случае имела место его адекватная пространственная фиксация и формирование перцептивного образа. Вместе с тем, то же правое пространство проецируется в височную зону не правого, а левого глаза. В соответствии с этим объект, который монокулярно предъявляется височной зоне левого глаза, в бинокулярном поле зрения оказывается справа (образ строится в данном случае правым глазом). Здесь имеет место его фантомный образ.
Характеризуя фантомные образы, мы выделили как одно их из основных свойств неустойчивость их к внешнему воздействию. Причиной такой неустойчивости может быть борьба полей зрения, которая неизбежна при проекции двух различных раздражителей в корреспондирующие зоны сетчаток. Так, если в фовеа одного глаза спроецировать какой-то раздражитель, то восприятие его будет конкурировать с восприятием раздражителя, не соответствующего ему по форме, цвету или величине, спроецированного в фовеа другого глаза. Аналогично этому при предъявлении какого-то раздражителя височной зоне одного глаза восприятие его будет конкурировать с восприятием другого раздражителя, предъявляемого носовой зоне второго глаза. Иначе говоря, борьба полей зрения имеет место в том случае, если искусственно создаются условия для разобщенного функционирования корреспондирующих зон как самостоятельных независимых систем. Тогда каждая из зон участвует в создании своего перцептивного образа, т. е. борьба полей зрения происходит при столкновении двух перцептивных образов.
Если с учетом этих механизмов рассмотреть наши опыты, то не трудно понять существенные различия между перцептивными и фантомными образами. В случае проекции раздражителя в височную зону одного глаза и появления движущегося объекта (указки) в пространстве носовой зоны другого имеет место несовпадающая стимуляция пары корреспондирующих зон, которая и вызывает борьбу полей зрения. Дополнительный движущийся раздражитель в этом случае оказывается конкурирующим с фантомным образом, который под его влиянием исчезает.
В случае проекции раздражителя в носовую зону (условие формирования перцептивного образа) появление движущейся указки в пространстве той же носовой зоны не приводит к борьбе полей зрения, так как дважды стимулируется только одна из пары корреспондирующих зон. Тот же дополнительный раздражитель, временно-разрушающий фантомный образ, не конкурирует с перцептивным образом: движение указки в зоне перцептивного образа не приводит к изменениям в его восприятии. Следовательно, неустойчивость фантомных образов, возникающих при проекции Стимулов в височные и фовеальные зоны сетчаток, можно рассматривать как показатель их восприятия в пространстве корреспондирующих зон. Вопрос только в том, почему раздражитель, занимающий совершенно определенную точку реального пространства, в большинстве случаев не воспринимается в этой точке (в условиях наших опытов)2, а воспринимается в другой точке пространства, связанной с первой функционально, т. е. почему вместо перцептивного образа формируется фантомный. Удивительным является и то, что перцептивный образ заменяется фантомным у людей с нормальным бинокулярным зрением и нормальной латерализацией церебральных функций. Вместе с тем мы имеем многочисленные экспериментальные данные, свидетельствующие о том, что у некоторых больных заиканием, в латерализации церебральных функций и билатеральной регуляции которых имеются значительные отклонения [4], [8] и др., в аналогичных условиях перцептивные образы не заменяются фантомными.
Рассматривая вопрос о природе фантомных образов, следует обратить внимание на то, что фантомные образы в периферическом зрении легче всего возникают при проекции раздражителей в височные зоны сетчатки (при отсутствии раздражителей в пространстве носовых зон) и при проекции раздражителей в одну из фовеальных зон (при отсутствии стимуляции фовеа другого глаза). Рецепторы именно этих зон имеют выходы к нейронам зрительной коры ипсилатеральных полушарий (неперекрещенные пути, соединяющие височные зоны периферического и центрального зрения, а также двойная проекция фовеальных зон в мозговые полушария).
В то же время проекция раздражителей в носовые зоны, рецепторы которых связаны по перекрещенным нервным путям с контралатеральиыми полушариями, так же как проекция в фовеальные зоны, тоже имеющие выходы к нейронам контралатеральных полушарий, в большинстве случаев приводит к возникновению не фантомных, а перцептивных образов. Здесь очень показательно, что раздражитель, проецируемый в носовые зоны, более жестко связанные с контралатеральными полушариями, значительно чаше вызывает появление перцептивного
121
образа (90% случаев), чем раздражитель, проецируемый в фовеальные зоны (в пределах 60% случаев), которые, видимо, изначально имеют равную возможность соединения с нейронами как ипсилатералыюго, так и контралатерального полушарий.
Появление фантомных образов в пространстве корреспондирующих зон позволяет предположить, что в этих случаях функциональная система двух корреспондирующих зон работает как открытая, незамкнутая система: получая зрительную информацию через одну корреспондирующую зону, мозг продуцирует фантомный образ в пространстве через вторую корреспондирующую зону. В соответствии с этим фантомный образ можно отнести к категории «визуализированных образов, т. е. образов, продуцируемых мозгом»3 [11].
В соответствии с этим определением предположение о репродуктивной природе фантомных образов можно обосновать не только их свойствами (положение в пространстве корреспондирующих зон и неустойчивость к внешнему воздействию), но и особенностями строения зрительного анализатора. Слой мозговой коры 17-го поля, куда приходит возбуждение по перекрещенным волокнам от рецепторов носовых зон периферического и центрального зрения (подслой "С" IV слоя), по густоте расположения, разнообразию форм и строения, сложности сплетений аксонных и дендритных ответвлений как будто предназначен для поточечной проекции сетчаточного возбуждения [29], [32], [33]. А слой IV "А", куда приходит возбуждение по неперекрещенным волокнам от височных половин периферического и центрального зрения, имеет совершенно другое строение, характерной особенностью которого является рыхлость сплетения афферентных волокон (там же). Но главный интерес представляет подслой IV "В", в котором происходит слияние возбуждений, приходящих из подслоев IV "А" и IV "С", и который считается анатомическим субстратом фузии [28], [32] и др. Этот слой содержит большие звездчатые клетки Кахаля, дающие начало эфферентной системе зрительного анализатора, связывающей нейроны коры 17-го поля с рецепторами сетчатки [15], [17], [22], [32] и др. Толстые быстропроводящие волокна эфферентных нейронов, начинающиеся от клеток Кахаля, прослеживаются на всех участках зрительного пути. В зрительном нерве они составляют около 10% [27; 111].
Тот факт, что эфферентный путь начинается в подслое IV "В", ответственном за фузионные процессы, не может не наталкивать на предположение об участии в актах бинокулярного зрения образов, репродуцируемых мозгом. Основываясь на указанных особенностях строения зрительного анализатора, также можно предположить участие образов, репродуцируемых мозгом, именно в фузионных процессах.
Следует обратить особое внимание и на то, что фантомные образы не локализовались в наших наблюдениях билатерально, а всегда смещались в контралатеральном направлении (по отношению к предъявляемому стимулу), что свидетельствует об их зависимости от деятельности нейронов именно 17-го проекционного поля, связанного исключительно с контралатеральным пространством [19], [20], [21] и др.
В отличие от визуализированных образов, связанных с содержательной стороной предметного зрения и опосредуемых деятельностью ассоциативных зон коры, фантомные образы, так же как и перцептивные, видимо, связаны с деятельностью проекционного, 17-го поля зрительной коры (ответственного за фузию). Именно этим, вероятно, объясняется одинаковая возможность продуцирования фантомных образов как центральным, так и периферическим зрением.
Судя по нашим экспериментальным данным и исходя из особенностей строения проекционного поля зрительной коры, можно предположить, что фантомный образ не артефакт бинокулярного зрения, но, наряду с перцептивным образом, является равноправным и необходимым компонентом фузионного образа. Видимо, в каждый момент только через один глаз, через рецепторы одной из корреспондирующих зон мозг адекватно фиксирует пространственное положение объекта, создавая его непосредственное чувственное впечатление (перцептивный образ). В то же время через второй глаз, через рецепторы второй корреспондирующей зоны мозг продуцирует фантомный образ воспринимаемого объекта. Оба эти процесса — чувственного впечатления (перцепции) и репродуцирования образа в естественных условиях бинокулярного зрения протекают как единый и неразрывный функциональный акт, результатом которого является единый продукт (фузионный образ).
Вопрос о значении визуализированного образа в актах восприятия ставился и ранее. Но другие исследователи ([12], [21] и др.) имели в виду не только вынесение в пространство образа, продуцируемого мозгом, но и соединение его с реальным объектом, тогда как мы говорим о соединении двух образов (перцептивного и фантомного).
На основе наших опытов можно предположить, что как перцептивные, так и фантомные образы могут обеспечиваться деятельностью любой из морфофункциональных систем бинокулярного зрения. Вместе с тем в периферическом бинокулярном зрении рецепторам носовых зон, связанным с нейронами контралатеральных полушарий, свойственно специализироваться на перцептивных процессах и создании перцептивных образов, тогда как ре-цеторам височных зон, имеющим проекцию на нейроны ипсилатеральных полушарий, свойственно специализироваться на репродуктивных процессах и создании фантомных образов. В центральном бинокулярном зрении участие фовеальных зон в создании фузионного образа также не одинаково — фовеальная зона правого глаза чаще обеспечивает появление перцептивного образа, а левого — фантомного образа. Причисляя фантомный образ к категории визуализированных образов, следует подчеркнуть, что и от визуализированных образов фантомные образы существенно отличаются.
Визуализированные образы составляют широкий круг зрительных образов, которые продуцируются мозгом чаще в состояниях, отличных
122
от спокойного бодрствования. В состояниях сна, гипноза, сенсорной депривации у человека могут возникать зрительные образы, которые в силу их яркости, четкости и динамичности воспринимаются как реальные объекты [7], [24), [30], [31]. Подобные гипнотические образы могут возникать при различных патологических состояниях мозга (в виде галлюцинаций), при непосредственном воздействии на структуры открытого мозга электрическим током («эффект присутствия» [25]) или вследствие фармакологического воздействия [3].
Кроме гипнотических к визуализированным образам, видимо, следует также отнести последовательные и эйдетические образы, широко изучаемые экспериментально ([7], [10], [26] и др.). В настоящее время делаются серьезные и, видимо, обоснованные заявки на возможность объективной регистрации визуализированных образов, в частности последовательных [26] и галлюцинаторных [18].
Все виды визуализированных образов как образов, продуцируемых мозгом, объединяет их временной отрыв от непосредственного восприятия и базирование на следах перцептивных процессов [13]. Фантомные образы, напротив, имеют место только в присутствии раздражителя, в момент его непосредственного восприятия. Переходя на язык психологических понятий, можно сказать, что фантомный образ включен в восприятие, является его необходимым компонентом. Репродуктивная природа фантомного образа как компонента фузионного образа позволяет нам проникнуть в сущность билатерального взаимодействия двух монокулярных систем, соединяющихся в одну бинокулярную. И в этом смысле какие-то аналоги фантомных образов зрительного восприятия не исключены и в деятельности анализаторов других модальностей, также имеющих двойной рецептор, парные мозговые центры и билатеральную регуляцию.
В отличие от визуализированных образов, занимающих в психике и поведении человека существенное место, особенно у людей первосигнального, художественного типа, фантомные образы имеют более ограниченное значение. Но чрезвычайно существенно то, что фантомные образы входят в интимные механизмы бинокулярного зрения и могут рассматриваться как модель организации психонервной деятельности человека.
Вопрос об участии в бинокулярном зрении фантомных образов как образов, продуцируемых мозгом, видимо, ставится впервые. Но постановка проблемы объективирования ощущений, в частности зрительных, в связи с наличием эффекторных систем в анализаторах имеет давнюю историю. Наиболее широко эта проблема поставлена В. М. Бехтеревым, который считал, что эфферентные импульсы в чувствующих органах «играют известную роль в объективировании получаемого ощущения, т. е. в отнесении его наружу» [6; 305]. «В самом деле,— писал он,— трудно было бы понять закон проекции наших ощущений, если бы в наших чувствующих проводниках и соответствующих им органах чувств не было условий для обратного движения нервного тока в периферическом направлении» (там же).
На тех же позициях стоял и. другой крупнейший невропатолог-клиницист, С. С. Корсаков: «Каждое наше конкретное представление вызывает хотя бы слабый центробежный иннервационный процесс, сопровождающийся слабым, но вполне соответствующим ему возбуждением периферических нервных элементов в органах чувств, что при каждом представлении о предмете мы хотя и в минимальной степени, но по-настоящему его видим или слышим, вообще ощущаем» [16; 48]. В дальнейшем на основе этих теоретических предпосылок И. С. Беритов разработал концепцию «образной психонервной деятельности». Он доказал экспериментальную роль образов, продуцируемых мозгом, в поведении животных и человека. Причем именно И. С. Беритов подробно обосновал зависимость возникновения зрительных образов от эфферентной системы зрительного анализатора и соотнес их проекцию в пространство с активностью нейронов ассоциативных зрительных полей (18 и 19) [5]. В настоящее время зрительные образы снова становятся объектом исследований психологов и физиологов ([7], [30], [31] и др.). Особенно привлекательной представляется проблема зрительной образной памяти в связи со значением ведущего анализатора в невербальном интеллекте и доминированием правого полушария. Наличие же фантомного образа как предтечи зрительного представления в таком, казалось бы, чисто перцептивном процессе, каким считается образование фузионного образа, может быть показателем значения билатеральной регуляции и межполушарной асимметрии для развития психических функций.
ВЫВОДЫ
1. В процессе изучения бинокулярного зрения как функции анализатора, имеющего парный рецептор и парные мозговые центры, удалось выявить и описать особый вид зрительных образов, названных фантомными.
2. Фантомный образ относится к категории образов, продуцируемых мозгом (визуализированных образов), и связывается с деятельностью эфферентной системы зрительного анализатора. В отличие от визуализированного образа фантомный формируется в процессе восприятия стимула и, наряду с перцептивным образом, является компонентом фузионного образа.
3. Фантомный образ выявляется при изолированной стимуляции одной из двух корреспондирующих зон центрального и периферического зрения и локализуется в пространстве, принадлежащем второй (не стимулируемой) корреспондирующей зоне.
4. Характерной особенностью фантомного образа является его неустойчивость к внешнему воздействию: движение дополнительного раздражителя в зоне видения фантомного образа вызывает его кратковременное исчезновение.
5. Следует предположить, что и в центральном, и в периферическом бинокулярном зрении появление фантомного образа обусловливается проекцией стимула в рецепторы, связанные с нейронами ипсилатеральных полушарий.
1. Ананьев Б. Г. Системный механизм восприятия пространства и парная работа полушарий
123
головного мозга. — В сб.: Проблемы восприятия пространства и пространственных отношений. М., 1961, с. 5—10.
2. Аветисов Э. С. Содружественное косоглазие. — М., 1977. —312 с.
3. Бабоян Д. Путевка в ад: Пер. с рум. — М., 1973.—232 с.
4. Бережковская Е. Л., Голод В. И., Туровская З. Г. Сенсорная асимметрия у здоровых и больных логоневрозом. — Вопросы психологии, 1980, № 1,с. 57—63.
5. Беритов И. С. Структура и функции коры большого мозга. — М., 1969, с. 70—100, с. 205— 231.
6. Бехтерев В. М. Проводящие пути спинного и головного мозга. Ч. 2. — СПб., 1898. — 305 с.
7. Браун Д. Послеобразы. — В кн.: Зрительные образы: феноменология и эксперимент. Ч. I.-Душанбе, 1972, с. 93—145.
8. Данилов И. В., Черепанов И. М. Патофизиология логоневрозов. — Л., 1970, с. 58— 127.
9. Джадд Д. Основные корреляты зрительного раздражителя. — В кн.: Экспериментальная психология, т. 2 / Под ред. С. С. Стивенса. М., 1963—377 с.
10. Зенкин Г. М., Петров А. П. О механизмах константности зрительного восприятия пространства. — В кн.: Сенсорные системы. Механизмы зрения. Л., 1979, с. 25—39.
11. Зинченко В. П., Вергилес Н. Ю. Формирование зрительного образа. — М., 1969.— 109 с.
12. Злоказов В. П., Пушкин В. Н., Шевчик Э. Д. Биоэнергетические аспекты соотношения образа восприятия с воспринимаемым объектом. — В сб.: Вопросы психогигиены, психофизиологии, социологии в угольной промышленности и психоэнергетики. М., 1980, с. 358— 368.
13. Клацки Р. Память человека. Структура и процессы. — М., 1978. — 319 с.
14. Коган А. И. Бинокулярная система и восприятие трехмерного пространства. — В кн.: Физиология сенсорных систем. Физиология зрения. Ч. I. Л., 1971, с. 279—303.
15. Кононова Е. П. Проводящие пути зрительных галлюцинаций. — Мат. 3-го конгресса по психотронике. — Япония, 1977, т. 2, с. 487—497 (на рус. яз.).
16. Корсаков С. С. Избранные произведения. — М., 1954.— 48 с.
17. Кравков С. В. Глаз и его работа. — М.; Д., 1950, с. 5—68.
18. Крохалев Г. П. Фотографирование зрительного анализатора. — В кн.: Анатомия и гистология нервной системы. М., 1957, с. 185— 190.
19. Куффлер С., Николе Дж. От нейрона к мозгу. — М., 1979, с. 26—67.
20. Лурия А. Р. Основы нейропсихологии. — М., 1973, с. 130—133.
21. Меграбян А. А. Общая психопатология. — М., 1972, с. 23.
22. Милнер П. Физиологическая психология. — М., 1973, с. 242—243.
23. Мэгун Г. Бодрствующий мозг. — М., 1961, с. 126.
24. Нейссер У. Зрительные процессы. — В кн.: Восприятие. Механизмы и модели. М., 1974, с. 250—261.
25. Пенфилд У., Джаспер Г. Эпилепсия и функциональная анатомия головного мозга. — М., 1958.
26. Сергеев Г. А. Об электромагнитном излучении плазменных пучков биологического происхождения нестационарной природы. — В сб.: Свет гелий-неоновых лазеров в биологии и медицине. Алма-Ата, 1970, с. 63—64.
27. Сутро Л. Модель зрительного пространства. — В кн.: Проблемы бионики. М., 1965, с. 107—126.
28. Трон Е. Ж. Заболевания зрительного пути. — М., 1968, с. 3—53.
29. Филимонов И. Н. Затылочная область. — В кн.: Анатомия и гистология нервной системы. М., 1957, с. 20—29.
30. Хебб Д. Об образах. — В кн.: Зрительные образы: феноменология и эксперимент. Ч. I. Душанбе, 1972, с. 145—164.
31. Хольт Р. Образы: возвращения и изгнания. — В кн.: Зрительные образы: феноменология и эксперимент. Ч. I. Душанбе, 1972, с. 51—72.
32. Школьник-Яррос Е. Г. Нейроны и межнейронные связи. Зрительный анализатор. — М., 1965. —227 с.
33. Школьник-Яррос Е. Г. Морфология зрительной системы. — В сб.: Физиология сенсорных систем. Физиология зрения. Ч. I. Л., 1971, с. 14—36.
34. Blakemore С, Pettigrew J. D. Eye dominance in the visual cortex. — Nature, 1970, v. 225, N 5231, p. 426—429.
35. Cogan A. J. Qualitative observations in visual science. The farm-sworth shelf. Fusion at the site of the "chosts". — Vision Res., 1978, 18, № 6, p. 657—664.
36. Nlkara Т., Bishop P. O., Pettigrew S. D. Analysis of retinal correspondence by studying receptive fields of binocular single units in the cat striat cortex. — Exp. Brain Res., 1968, v. 6, p. 353-372.
Поступила в редакцию 05.I.1982 г.
1 В дальнейшем пространство, проецируемое в определенную зону сетчатки, мы будем условно называть пространством, принадлежащим этой зоне.
2 Аналогичные экспериментальные результаты были получены и в реальном пространстве не ограниченном экраном [35].
3 Под визуализированными образами мы понимаем широкий круг зрительных образов, репродуцируемых мозгом на основе следов перцептивных процессов, т. е. образы зрительных представлений.