83
К ВОПРОСУ
О ПСИХОФИЗИОЛОГИЧЕСКИХ МЕХАНИЗМАХ
ПРОИЗВОЛЬНОГО ВОСПРОИЗВЕДЕНИЯ
Я. В. БОЛЬШУНОВ
Новосибирский педагогический институт
Настоящее исследование посвящено вопросу о психофизиологических механизмах избирательности и реконструктивности преднамеренного воспроизведения. С нейродинамической точки зрения, произвольная регуляция психической, деятельности рассматривается обычно как, второсигнально опосредствованный процесс [5], [6], [12], [18], [19], однако применительно к преднамеренному воспроизведению это положение нуждается в специальной разработке и конкретизации. Согласно поддерживаемой нами гипотезе, в процессе реконструктивного избирательного припоминания происходит селективная активация и направленная перестройка ранее образованных временных нервных связей в соответствии с реализуемой человеком репродуктивной задачей. В основе этого предположения лежит представление о механизме второсигнальных управляющих импульсов [5], [6], [17], [18], [19].
МЕТОДИКА
Испытуемому дается инструкция как можно быстрее двигательно реагировать на зрительно предъявляемые слова. При этом надо учитывать их значение. Если предъявляется название части тела, например «рука», то нажимать на кнопку нельзя, во всех же остальных случаях требуется максимально быстрый двигательный ответ. Сначала испытуемый учится дифференцированно реагировать на названия частей тела и любые другие слова. Обучающий этап имеет место во всех экспериментальных сериях. После этого опыт с реакциями временно прерывается и вводится задача на заучивание.
Испытуемому предлагаются для запоминания 22 слова на карточках (по одному на каждой). Половина этих слов обозначает цветы, остальные — названия городов СССР. Испытуемый раскладывает материал на две смысловые группы и заучивает его. Затем возобновляется эксперимент с реакциями. Испытуемый получает инструкцию: «Через окошко прибора будут предъявляться различные слова. Среди них имеются и взятые из карточек, и новые. Необходимо реагировать как можно быстрее на все слова, кроме тех, которые обозначают части тела. И еще обязательное условие. По ходу опыта будут ставиться задачи вспомнить то одно, то другое слово из карточек. Например, потребуется воспроизвести название какого-то города или цветика, которое начинается и оканчивается определенными буквами. Вам нужно сказать соответствующее слово вслух и держать его в уме до тех пор, пока не будет поставлена новая задача. Просьба точно выполнять это условие. В то же время помните, что нужно максимально быстро и правильно реагировать на предъявляемые слова».
Проведены две основных серии опытов. В Г ставились задачи вспоминать названия городов, во II — цветов. По ходу репродуктивной деятельности испытуемому предъявляются через рефлексометр слова из списка, в том числе каждый раз то, которое он в данный момент активно воспроизводит. Двигательный ответ на этот вербальный стимул рассматривается как тест-реакция, характеризующая функциональное состояние актуализированного мнемического следа. Например, дается инструкция: «Вспомните название цветка, которое начинается и оканчивается буквами «ф» и «а». Назовите вслух. Держите в уме». Испытуемый воспроизводит слово «фиалка» и акцентирует на нем свое внимание. Одновременно с выполнением этого
84
задания он должен быстро и правильно реагировать на вербальные стимулы, которые последовательно появляются в окошке рефлексометра (в данном случае: платок, фиалка, Ереван, плечо). Затем ставится задача на воспроизведение и удержание в уме названия другого цветка и т. д.
Помимо двух основных, проведена контрольная серия опытов. В ней использовался тот же список вербальных стимулов, однако не было ни предварительного заучивания 22 слов, ни постановки задач на воспроизведение. Цель контрольного эксперимента состояла в том, чтобы сопоставить латентные периоды реагирования на названия городов и цветов в условиях отсутствия специально направленной мнемической и репродуктивной деятельности. Исследование проведено на .30 студентах. Во всех сериях участвовали разные испытуемые (по 10 человек в каждой).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Статистические показатели результатов исследования представлены в табл. 1. В ней сопоставляется латентное время реагирования (ВР) на вербальные стимулы в двух основных и контрольной сериях опытов. Каждое указанное в таблице среднее значение имеет в своей основе 110 замеров.
Таблица 1
ДИНАМИКА ВР (МС) НА НАЗВАНИЯ ГОРОДОВ И ЦВЕТОВ В ЗАВИСИМОСТИ
ОТ РЕПРОДУКТИВНЫХ ЗАДАЧ
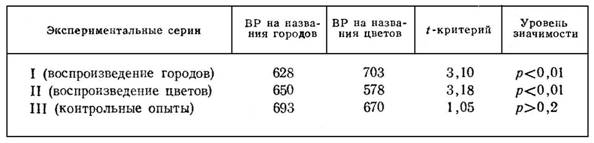
Как показывают полученные данные, в контрольном эксперименте сравниваемые показатели не отличаются значимо друг от друга. Иные результаты получены в основных опытах, в которых задача на реагирование выполнялась по ходу специально организованной репродуктивной деятельности. В этих условиях латентное время двигательных ответов в среднем заметно меньше на слова, вовлеченные в процесс произвольного воспроизведения. Испытуемые быстрее реагируют на названия городов (I серия) или цветов (II серия) в зависимости от того, что именно они активно вспоминают и удерживают в уме.
Таблица 2
ДИНАМИКА ВР НА СЛОВА «КИЕВ» и «РОЗА» (МС)
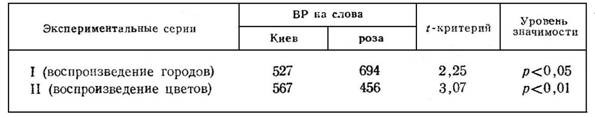
Заслуживает специального внимания динамика ВР на слова «Киев» и «роза» (табл. 2). Они имеют некоторые сходные особенности, от которых зависит быстрота двигательных ответов. Оба слова предъявляются в конце эксперимента, когда испытуемые уже полностью адаптированы к его условиям, причем следуют непосредственно друг за другом. Они одинаковы по «длине» (в каждом 2 слога и 4 буквы). Кроме того, они достаточно хорошо представлены в речевом опыте студентов. Поэтому сравнение ВР на слова «Киев» и «роза», по-видимому, особенно показательно.
85
Сопоставляемые в табл. 2 средние арифметические имеют в основе по 10 замеров. Оценка достоверности их различия производится по t-критерию для малых выборок, способ вычисления которого рассматривается в работе Э. Колкот [9].
Результаты, представленные в табл. 2, ясно говорят о том, что ВР на каждое слово существенно определяется постановкой соответствующих репродуктивных задач. Испытуемые дают более быстрые двигательные ответы на тот вербальный стимул, который согласно инструкции вовлекается в процесс произвольного воспроизведения. Поэтому в I и II экспериментальных сериях происходит противоположно направленное изменение показателей скорости реагирования на слова «Киев» и «роза».
Итак, более высокая степень готовности к двигательным ответам обнаруживается в отношении вербальных стимулов, вовлеченных в репродуктивное действие. Это явление неодинаково выступает в отдельных опытах. В некоторых случаях оно наблюдается очень четко. Например, у испытуемой К. Н. в I серии эксперимента ВР на названия городов и цветов составляет в среднем соответственно 534 и 691 мс. Критерий достоверности различия этих показателей равен 2,25 (р<0,05). Перед испытуемой Б. О. ставятся задачи на воспроизведение слов, обозначающих цветы (II серия). Ее двигательные реакции на эти вербальные стимулы имеют среднее латентное время 492 мс, тогда как на названия городов — 680 мс (t = 3,43; p< 0,01). Приведенные примеры не единичны.
Вместе с тем в некоторых опытах рассматриваемое явление выражено слабо или вообще не наблюдается, причем индивидуальные различия выступают весьма отчетливо. У 13 человек (из 20, участвовавших в основных экспериментальных сериях) усредненное латентное время двигательных ответов короче в пределах от 36 до 284 мс на слова, которые вовлечены в репродуктивный процесс. У испытуемой М. О. наблюдается обратное явление. Она заметно медленнее, хотя и на незначимом уровне (t = 1,27; р>0,2), реагирует на вербальные стимулы, подлежащие воспроизведению и удержанию в уме. Имеются испытуемые, у которых среднее латентное время двигательных ответов на. названия городов и цветов практически одинаково.
Таким образом, соотношение скорости реагирования на слова, включенные и не включенные в репродуктивный процесс, не во всех опытах однозначно. Причины наблюдаемого варьирования показателей пока недостаточно ясны и заслуживают специального изучения. Возможно, одна из них заключается в том, что не все испытуемые полностью соблюдают данную им инструкцию. Некоторые, судя по их словесным отчетам, начинают по собственной инициативе мысленно вспоминать не только требуемые слова, но и входящие во вторую группу вербальных стимулов или же настраивают себя на предугадывание моментов, когда будет предъявлено очередное название части тела. Очевидно, что такого рода самоинструкции могут существенно влиять на результаты опытов. Как отмечает О. А. Конопкин [10], то или иное понимание субъектом экспериментальной задачи и специфическое самопрограммирование им своей деятельности являются важными психологическими механизмами регуляции сенсомоторных актов. Однако, несмотря на межиндивидуальные различия, в целом по каждой основной серии опытов двигательные реакции осуществляются достоверно быстрее на слова, которые одновременно вовлечены в процесс произвольного воспроизведения.
Каков психофизиологический смысл этого явления? Многие исследователи рассматривают ВР в качестве важного показателя готовности
86
к функционированию соответствующих нервных связей [5], [14], [17], [18]. Так, Е. И. Бойко пишет: «По изменениям скрытого периода пробной реакции можно следить за состоянием возбудимости зрительно-двигательных второсигнальных путей в зависимости от того, какое одиночное слово является тестирующим» [6; 123]. В проведенных опытах каждое название города или цветка предъявляется по ходу выполнения определенного репродуктивного действия. При этом обнаруживается достоверное различие в скорости реагирования на слова, вовлеченные и невовлеченные в процесс произвольного воспроизведения. Результаты эксперимента позволяют говорить о том, что в основных сериях опытов динамика латентных периодов двигательных ответов на названия городов и цветов в значительной мере определяется функциональным состоянием тестируемых следов памяти. Тем самым получает подтверждение гипотеза о важной роли механизма второсигнальных управляющих импульсов в регуляции произвольного воспроизведения. Их источником является, по-видимому, мозговая энграмма репродуктивного намерения, возникающая под влиянием определенного запроса к памяти человека. Второсигнальные положительные импульсы активируют следы прошлого опыта, ассоциированные с признаками требуемого воспроизведения. В результате происходит избирательное повышение готовности к функционированию тех временных нервных связей, которые соответствуют стоящей перед субъектом репродуктивной задаче.
Развиваемая концепция произвольного воспроизведения согласуется с фактами, полученными в электрофизиологических исследованиях мнемической деятельности человека. В работах М. Н. Ливанова и С. Н. Раевой [11], а также С. Н. Раевой [15] показано, что существуют особые нейрональные реакции, возникающие в ответ на задания типа «запомните!», «повторите!», «приготовьтесь!» и т. д. Авторы отмечают, что словесные стимулы такого рода имеют значение как бы «пусковых» сигналов, предваряющих выполнение поведенческих (мыслительных или двигательных) актов. Специфическое функционирование этого типа нейронов наблюдается, в частности, при проверке произвольной памяти с помощью теста Бине. Причем свойственный им фазический, «залповый» характер реагирования отчетливо выступает в начале мнемического действия и затем при переходе к воспроизведению материала. М. Н. Ливанов и С. Н. Раева считают, что специализированные ответы выделенного ими типа клеток человеческого мозга отражают процессы межнейронных перестроек, связанные «...скорее всего с механизмом, экстренной активации» [11; 23].
Как известно, различаются положительные и отрицательные (тормозящие) второсигнальные управляющие импульсы. Первые физиологически облегчают осуществление сенсомоторных актов, вторые, наоборот, задерживают, блокируют их, если они по условиям предварительного словесного задания оказываются ненужными или «запретными».
В регуляции репродуктивного действия участвуют, по всей вероятности, не только положительные, но и отрицательные управляющие нервные импульсы. Произвольное воспроизведение всегда направлено на актуализацию вполне определенного содержания прошлого опыта. Человек сознательно отграничивает нужные образы и мысли от тех, которые не соответствуют репродуктивной задаче. Посторонние, не относящиеся к делу ассоциации задерживаются. Причем блокирование не исчерпывается тем, что отбрасывается какое-то уже актуализированное в уме содержание прошлого опыта. Исследования [7], [8] показывают, что репродуктивная задача влияет на стратегию самого поиска в процессе воспроизведения. Например, испытуемый не стремится вспомнить какие-то частности, подробности воспринятого текста, если
87
предварительная инструкция нацеливает на изложение только основного содержания материала.
Физиологической основой блокирования в процессе произвольного воспроизведения является, по-видимому, механизм тормозящих второсигнальных управляющих импульсов, генерируемых энграммой репродуктивного намерения. Имеются факты, которые косвенно (на материале опознания слов) подтверждают это предположение.
Как указывалось, во всех трех рассмотренных экспериментальных сериях испытуемым запрещалось реагировать на названия частей тела. Это требование обеспечивало осознанное восприятие каждого предъявляемого через рефлексометр слова. Тем самым устранялись машинальные двигательные реакции на вербальные стимулы.
Однако практическая реализация «тормозного» предписания предварительной инструкции дается испытуемым нелегко, причем наблюдаются трудности двоякого рода. Во-первых, несмотря на стремление испытуемых точно выполнять поставленную задачу, они нередко дают «запретные» двигательные ответы на названия частей тела. Такие ошибки (от 1 до 3) встречаются у 24 человек из всех 30, участвовавших в эксперименте. Во-вторых, «тормозной» компонент инструкции может оказывать отрицательное влияние на опознание положительных вербальных стимулов. Иногда предъявляемое через рефлексометр слово неправильно воспринимается в первый момент как название какой-то части тела. Это ведет к задержке нужной реакции. Такое кратковременное неточное восприятие слов наблюдается в том случае, когда они по своему буквенному составу более или менее совпадают с названия ми частей тела. Данный факт привлек внимание уже в пробных опытах. Затем он стал предметом специального изучения. С этой целью в списки вербальных стимулов были включены сходные по буквенному составу слова «темя» и «тема». Первое предъявлялось в обучающей части эксперимента, второе — во время выполнения основного задания.
Полученные результаты свидетельствуют, что положительный вербальный стимул «тема» многими испытуемыми воспринимается сначала как «темя». В связи с этим резко задерживаются соответствующие моторные реакции. Латентное время двигательных ответов на слово «тема» в среднем значительно больше, чем на любой другой использованный в опытах положительный вербальный стимул.
Для сравнения целесообразно взять показатели скорости реагирования на смежные в списке и одинаковые по «длине» слова «Киев» и «роза» (табл. 3). Очевидно, что сопоставление следует проводить по результатам контрольного эксперимента, в котором названия городов и цветов не были объектом ни предварительного заучивания, ни выборочного воспроизведения. В этой серии все слова, в том числе «Киев», «роза», «тема», предъявляются испытуемым неожиданно. Каждая приведенная в табл. 3 средняя арифметическая основывается на 10 замерах.
Таблица 3
ВР НА СЛОВА «КИЕВ», «РОЗА», «ТЕМА» (М)
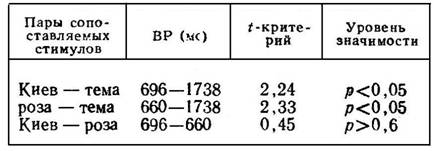
Из табл. 3 видно, что показатели скорости реагирования на стимулы «Киев» а «роза» различаются на незначимом уровне. Особое положение занимает слово «тема». Среднее латентное время двигательных ответов на его предъявление в два с половиной раза больше, чем на другие вербальные стимулы.
88
Этот факт явно связан с влиянием «тормозного» компонента предварительной инструкции. Испытуемые, стремясь избежать ошибочных ответов на названия частей тела, настраиваются в той или иной степени на ожидание соответствующих вербальных стимулов. Поэтому во многих случаях слово «тема» опознается первоначально как ранее предъявленное «темя». Оно воспринимается на какой-то момент в качестве «запретного» для реагирования. В связи с этим приводится в действие механизм второсигнальных тормозящих импульсов. В результате задерживаются двигательные реакции на слово «тема».
С избирательностью произвольного воспроизведения тесно связан процесс реконструкции актуализируемого материала. Известно, что, припоминая давнее событие или ранее воспринятый текст, человек более или менее перестраивает его. Обширный материал по этому вопросу содержится в исследованиях Ф. Бартлетта [1], П. П. Блонского [4], А. А. Смирнова [16] и других психологов. В. Я. Ляудис [13] характеризует реконструктивное избирательное воспроизведение как оптимальный уровень готовности памяти к актуализации нужной информации, который возможен только при достаточно высокой сформированности соответствующего умственного действия.
Особенно большое влияние на реконструкцию актуализируемых образов и мыслей оказывают специальные репродуктивные задачи. Например, в зависимости от того, требуется ли изложить воспринятый текст подробно, полностью или, наоборот, кратко, только его основное содержание, воспроизведение значительно изменяется как по объему, так и по видам перестройки [7]. Это означает, что оно не предопределяется жестко теми временными нервными связями, какие существуют в мозгу человека до начала репродуктивного действия. Обобщение смысловых звеньев вспоминаемого текста, объединение и дробление его отдельных частей, изменение последовательности, привнесение дополнительного содержания имеют своей физиологической основой, по всей вероятности, не просто оживление прежних нервных следов, но их определенное преобразование. А. А. Смирнов отмечает, что перестройка актуализируемого материала иногда настолько значительна, что «...сам процесс воспроизведения в этих случаях никак не может рассматриваться как голая репродукция, как простое восстановление того, что было воспринято» [16; 157].
Реконструкция в процессе припоминания текста означает по своей психофизиологической природе какие-то изменения ранее сложившихся нервных следов. Она происходит, по-видимому, благодаря особого рода временным связям, которые Е. И. Бойко называет динамическими. Их специфика состоит в том, что они экстренно создаются через вторую сигнальную систему, возникая как продукт текущей нейродинамики в процессе переработки прошлого опыта. Е. И. Бойко пишет, что они, в отличие от замыкательных (ассоциативных) связей, представляют собой «...экстренные преобразования действующих нервных путей по ходу уже начавшейся сложной реакции» [6; 241],
Очевидно, что концепция простого оживления следов памяти не позволяет удовлетворительно объяснить направленную перестройку актуализируемого материала, обусловленную тем или иным специальным репродуктивным намерением субъекта. Более приемлема гипотеза, согласно которой реконструированное произвольное воспроизведение имеет своей физиологической основой слаженную систему замыкательных и динамических временных связей. Е. И. Бойко обозначает этот механизм умственной деятельности человека термином «динамическая вариотипия» [6; 190—191].
Каждый вариотип включает в себя как относительно устойчивые, сложившиеся в прошлом опыте, так и экстренно формирующиеся нервные
89
пути. Причем эта система замыкательных и динамических временных связей складывается под влиянием предварительного словесного задания. Применительно к произвольному реконструированному воспроизведению в ней реализуется конкретное репродуктивное намерение субъекта. Иначе говоря, поиск, восстановление и переструктурирование нужных следов памяти происходит по механизму второсигнальных управляющих импульсов.
В связи с обсуждаемой гипотезой большой интерес представляют нейрофизиологические исследования И. П. Бехтеревой [2], [3]. Она исходит из того, что в мозговых системах обеспечения психической деятельности имеются как «жесткие», сложившиеся в прошлом опыте, так и «гибкие», экстренно включающиеся компоненты. Благодаря последним поведение человека организуется с учетом изменяющейся внешней и внутренней среды организма. На основе экспериментального материала Н. П. Бехтерева заключает, что в условиях выполнения психологических тестов, в том числе на кратковременную память, «...формируются рабочие нейронные ансамбли, в данном случае функционально объединенные в соответствии с реализуемой задачей» [2; 105].
Рассмотренные факты позволяют сделать вывод, что то или иное специальное репродуктивное намерение может вести к преобразованию следов прошлого опыта. В мозговую систему обеспечения произвольного реконструктивного воспроизведения входят, по-видимому, различные по «жесткости» — «гибкости» звенья. В совокупности они составляют вариотип временных нервных связей, формирующийся под влиянием стоящей перед человеком репродуктивной задачи.
1. Бартлетт Ф. Психика человека в труде и игре.— М., 1959.— 144 с.
2. Бехтерева Н. П. Нейрофизиологические аспекты психической деятельности человека. — 2-е изд.— Л., 1974. — 151 с.
3. Бехтерева Н. П. Роль индивидуально приобретенной памяти в механизмах нормальных и патологических реакций. — В кн.: Механизмы модуляции памяти. Л., 1976, с. 7—14.
4. Блонский П. П. Избранные психологические произведения. — М., 1964. — 547 с
5. Бойко Е. И. Время реакции человека. — М., 1964. — 440 с.
6. Бойко Е. И. Механизмы умственной деятельности. (Динамические временные связи). — М., 1976. —248 с.
7. Большунов Я. В. Влияние специальных задач на процесс произвольного воспроизведения.— В кн.: Память и ее развитие. Ульяновск, 1973, с. 5—25.
8. Большунов Я. В. К характеристике структуры произвольного воспроизведения.— Вопросы психологии, 1978, № 3, с. 91—97.
9. Колкот Э. Проверка значимости. — М., 1978. — 128 с.
10. Конопкип О. А. О психологической саморегуляции сенсомоторной деятельности.— В кн.: Проблемы общей, возрастной и педагогической психологии. М., 1978, с. 78—92.
11. Ливанов М. Н., Раева С. Н. Микроэлектродное изучение нейроиальных механизмов произвольной мнестической деятельности человека. — В кн.: Механизмы модуляции памяти. Л., 1976, с. 14—24..
12. Лубовский В. И. Развитие словесной регуляции у детей. — М., 1978. — 224 с.
13. Ляудис В. Я. Память в процессе развития.— М., 1976. — 255 с.
14. Рабинович М. Я. Нейронные механизмы условного рефлекса. — В кн.: Физиология высшей нервной деятельности. Ч. 2. М., 1971, с. 3—33.
15. Раева С. Н. О некоторых нейроиальных механизмах произвольной мнестической деятельности человека.— Доклады Академии наук СССР, 1975, т. 221, Мб, с. 1477—1480.
16. Смирнов А. А. Проблемы психологии памяти. — М., 1966. — 422 с.
1.7. Ушакова Т. Н. Функциональные структуры второй сигнальной системы. — М„ 1979.—248 с.
18. Чуприкова Н. И, Слово как фактор управления в высшей нервной деятельности человека. — М., 1967. — 327 с.
19. Чуприкова Н. И. Проблема детерминации поведения человека и некоторые психофизиологические механизмы второй сигнальной системы. — В кн.: Проблемы общей, возрастной и педагогической психологии. — М., 1978, с. 153—166.